Quando si pensa all’evoluzione degli organismi si cade nell’errore più banale di immaginarlo come un processo che avviene lungo una linea, dove gli animali e le piante si migliorano per piccole modificazioni graduali, generazione dopo generazione, dai più primitivi fino a quelli più evoluti. Per esempio, per quanto riguarda l’evoluzione delle piante si immagina spesso che proceda con le seguenti tappe:
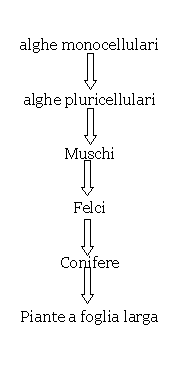
In realtà sarebbe più corretto immaginare l’evoluzione come un cespuglio fitto di rami (fig. 1 e 2).
fig. 1
fig. 2
Infatti, alcuni studiosi ritengono che le felci derivino direttamente dalle alghe, di conseguenza muschi e felci si sarebbero evoluti parallelamente giungendo fino a noi su strade diverse. Secondo altri, invece, la rassomiglianza delle felci nei primissimi stadi di sviluppo con un gruppo collaterale ai muschi (le Epatiche) sarebbero fin troppo evidenti. Resta aperta, dunque, la questione se le piante superiori derivino dai muschi, dalle epatiche o dalle alghe.
E’ certo comunque che tutti e tre i tipi di organismi abbiano avuto un progenitore comune a vita acquatica, una particolare specie di alga.
Le relazioni di parentela tra i diversi gruppi di alghe conosciuti, sia fossili che viventi, ci sono ancora oscuri. In ogni caso è probabile che da un gruppo di alghe denominato Cianoficee – considerate le più primitive in quanto costituite da un‘unica cellula come i batteri, prive di organi di propulsione (flagelli), di nucleo, di cloroplasti, ma in grado di liberare ossigeno gassoso – prese origine un gruppo leggermente più evoluto per la presenza di flagelli: le Euglenali. Da loro si svilupparono le Clorofite, più complesse per la presenza di una parete cellulare: queste sono risultate particolarmente interessanti perché un sotto gruppo, quello delle Caroficee (del quale fanno parte i generi viventi come Chara e Nitella, fig. 3), sono considerate i capostipiti dei muschi e delle epatiche (Briofite), le prime vere piante terrestri. Infatti, i loro fusti primordiali (cauloidi) presentano già nodi, internodi e cellule apicali meristematiche (si veda la risposta http://www.vialattea.net/esperti/php/risposta.php?num=12336) come nei muschi e le piante superiori. Avevano riproduzione sessuata, erano in grado di fissarsi al fondo del mare con delle strutture simili a delle radici (rizoidi) e possedevano già delle ramificazioni e parti laminari simile e foglie (filloidi).
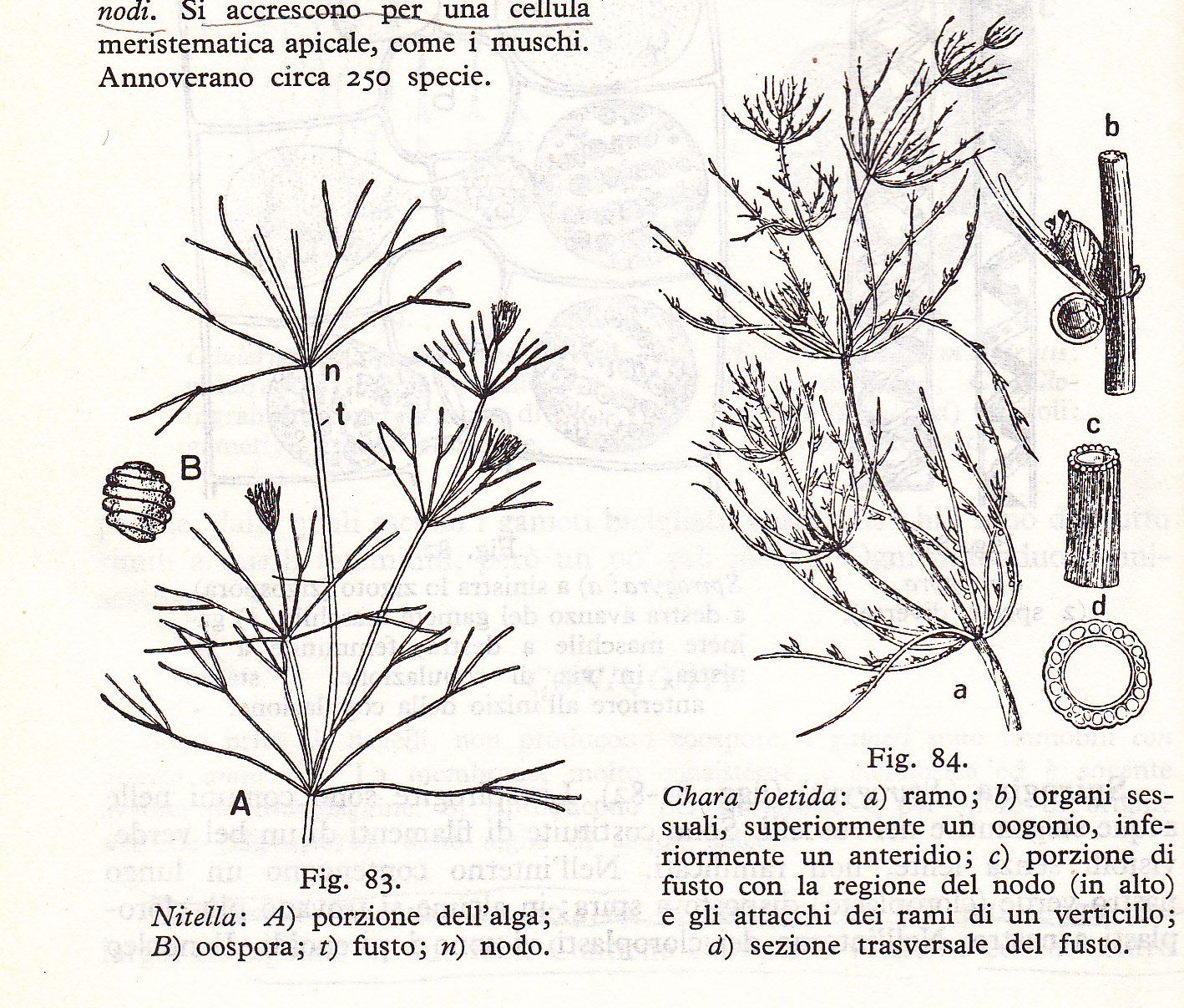
fig. 3
Le briofite più semplici (Marcanziali del gruppo delle epatiche) sono costituite da una semplice lamina (tallo) e appaiono come un nastro sdraiato, mentre le più complesse (Muschi) presentano già un fusticino eretto e foglioline ma non vere e proprie radici. Tra i due gruppi ve ne è uno intermedio, il Jungermanniali: presentano un tallo laminare ma più frequentemente è rappresentato da un fusticino sdraiato nel primo tratto e munito di rizoidi, poi diventa verticale con delle foglioline. Tra le epatiche esiste un gruppo assai evoluto e noto come Antocerotali che possono essere considerate il punto di origine delle felci, in particolare di un sotto gruppo (le psilofite) per la presenza di un maggior numero di tessuti clorofilliani e per il sistema di riproduzione.
Le alghe come i primi muschi, tuttavia, non possedevano un sistema di trasporto interno dei soluti (vasi conduttori); ogni singola cellula produceva il proprio nutrimento e aveva bisogno di essere in contatto con l’acqua. In realtà, un vero e proprio sistema di vasi di trasporto dei soluti si ebbe solo quando alcuni gruppi di alghe e briofite più audaci si avventurarono sulla terra ferma. In effetti, i geologi sostengono che nel Siluriano superiore (circa 415 milioni di anni fa) fossero presenti estese pianure di marea dove prosperavano particolari tipi di piante acquatiche capaci di resistere per brevi periodi all’aria. In alcuni casi i vantaggi in termini di sopravvivenza a questa condizione di vita aerea si rivelarono superiori ai costi. Infatti, per la prima volta era possibile ricavare maggiori quantitativi energia solare e anidride carbonica essenziali per la fotosintesi. Tuttavia, furono necessari alcuni adattamenti evolutivi. Il problema principale consisteva nel cercare di evitare l’eccesiva evaporazione e la conseguente disidratazione poiché i tessuti delle piante restavano a diretto contatto con l’aria e il calore del sole. Nelle prime Marcanziali per esempio, questo problema non sussisteva ancora poichè, l’ampio contatto con l’umidità del terreno, non solo facilitava l’assorbimento dei soluti, ma diminuiva anche i fenomeni di traspirazione. Nelle forme erette, invece, come i muschi più progrediti e a forma di piccoli alberi, la traspirazione così come la superficie fotosintetica risultava decisamente più elevata. Queste primissime piante terrestri allora evolsero un sistema per portare con sé un piccolo pezzetto di mare al loro interno. I rizoidi che si erano evoluti in un primo tempo per farle ancorare al fondo del mare, svilupparono una nuova funzione: assorbire l’acqua e i soluti dal terreno. A livello del cauloide e dei filloidi, invece, vennero premiate le cellule più esterne che si ispessivano per formare una sorta di barriera, la prima cuticola della storia che impediva l’evaporazione dei liquidi interni ma li regolava mediante delle piccole aperture, i cosiddetti stomi. Per portare l’acqua e i soluti dalle radici alle parti più alte del ‘fusto’, e gli zuccheri in direzione opposta, si sviluppò un sistema di vasi conduttori formato da cellule allungate. Le piante con queste caratteristiche di trasposto dei fluidi sono chiamate Cormofite.
La più antica pianta fossilizzata dotata di vasi è stata rinvenuta in Inghilterra: si tratta della Cooksonia caledonia (fig. 4); era alta circa 6 cm, ogni ramo a un certo punto si biforcava (dicòtimia) e all’apice era presente una capsula contenente le spore (sporangio); si è estinta senza lasciare discendenza.

fig. 4
Un altro fossile ancora più interessante dello stesso periodo è Rhynia (fig. 5), una pianta alta circa 17 cm somigliante ad una canna di bambù, viveva in ambienti paludosi; anch’essa possedeva rami dicotomici con sporangi. Dal gruppo a cui appartiene questo fossile (Rhyniophyta), successivamente, durante il Devoniano medio (circa 390 milioni di anni fa) sarebbe discesa Psilophyton dawsonii (fig. 6); questa psilofita primitiva sarebbe la diretta progenitrice delle attuali felci, conifere ed equiseti, tuttavia il suo modo di riprodursi era ancora vicino a quello delle briofite. Dall’asse principale di questa pianta si originano dei rami laterali, i quali a loro volta si biforcano più volte e sulla sommità di alcuni è possibile trovare gli sporangi. Le foglie sono ancora assenti, tuttavia in un altro fossile, Axteroxhylon (fig. 7), il fusto non appare liscio ma ricoperto di numerose foglioline (microfilli) disposte in modo embricato (come le tegole dei tetti), e senza dubbio aumentavano la superficie adibita alla fotosintesi. Secondo alcuni botanici va collocata nel gruppo delle felci anche se era ancora priva di vere e proprie radici ma queste, in ogni caso, erano ancora più evolute dei rizoidi poiché iniziavano a possedere i primi peli radicali impiegati per migliorare l’assorbimento dei soluti.

fig. 5
fig. 6

fig. 7
Dal gruppo delle psilofite si evolveranno nel Devoniano i tre sotto gruppi principali delle felci: Licopsida, Sfenopsida e Pteropsida. Il primo si caratterizza perché i suoi generi presentano fusti microfillacei e sono i diretti progenitori degli attuali Licopodi presenti nei boschi e nei pascoli montani. Grazie a un fossile risalente ad un periodo compreso tra 395 e 375 milioni di anni fa (Devoniano inferiore) che si chiamava Zosterophyllum, si è visto che queste piante dipendevano ancora dall’acqua per la riproduzione (zone d’estuario, di palude). Se potessimo tornare indietro a quell’epoca e visitare una zona paludosa, probabilmente incontreremo soprattutto vegetazione a livello erbaceo con molte piante appartenenti al genere Licopodium. Solo nell’epoca geologica successiva (Carbonifero: da 359 a 299 milioni di anni fa) potremmo inoltrarci veramente nella prima foresta preistorica con esemplari di Licopodi giganti (40 metri) come Lepidodendron (fig. 8) e Sigillaria che purtroppo si estinsero.

fig. 8
Queste piante, sebbene avessero un portamento di tipo arboreo, in realtà erano ancora erbacee; infatti, frequentemente venivano abbattute dal vento perché solo la parte più esterna del tronco era sostenuto da tessuto di consistenza legnosa e le radici si espandevano perlopiù in superficie. Il progressivo accumulo dei resti di queste prime foreste, insieme alla sedimentazione e costipazione, ha dato origine nel corso del tempo ha importanti depositi carboniferi.
Anche le Sfenopsida (o equiseti) risalgono al Devoniano come le Licopsida e raggiunsero la massima diffusione circa 300 milioni di anni fa, con individui che sfioravano altezze di 18 metri. La forma di queste piante, essendo primitive, seguiva criteri geometrici piuttosto semplici: la chioma difatti aveva forma conica per consentire anche ai rami e alle foglie disposte alla base del tronco di ricevere la luce che altrimenti si sarebbe attenuata dalla copertura fogliare soprastante (fig. 9). Come gli attuali equiseti il fusto era diviso in numerosi segmenti da varie costrizioni (nodi) in corrispondenza dei quali si dipartivano i rami e le foglie.

fig. 10
Le Pteropsida presentavano caratteri intermedi con le Psilofite e le attuali felci, e rispetto ai due gruppi precedenti, durante il carbonifero, erano meno numerose con individui di altezze inferiori, più spesso occupavano lo strato erbaceo. Il fossile Psaronius (fig. 11) mostra una pianta alta circa 3 metri con un tronco molto ampio alla base che si assottigliava verso l’alto, terminando con un ciuffo di fronde. Le foglie iniziavano ad avere la nervatura ramificata ed erano molto simili a quelle delle felci attuali (fig. 12).

fig. 11

fig. 12
Il punto di svolta, tuttavia, che segnò la conquista definitiva della terra ferma da parte delle piante fu l’evoluzione del seme. In pratica, la novità consiste nella possibilità del gametofito maschile (granulo pollinico) di raggiungere l’ovocellula femminile passando attraverso l’aria, lasciandosi trasportare dal vento. Inoltre, in questo modo la fecondazione avviene in modo più sicuro. Dopo la fecondazione, i rivestimenti cellulari del nuovo embrione cominciano ad ispessirsi fornendogli la possibilità di germinare solo al momento opportuno, nutrendolo quando le condizioni ambientali diventeranno favorevoli per lo sviluppo della piantina. L’ovocellula, quindi, con questi nuovi accorgimenti, diventa un seme, un organo dormiente in grado di risvegliasi e germinare dopo molto tempo, e soprattutto eventualmente lontano da chi lo ha generato, condizione essenziale per evitare la competizione e colonizzare nuovi territori. Prima dell’avvento del seme, le spore per poter germinare e trovare il corrispettivo gamete di sesso opposto, dipendevano ancora dall’acqua. Senza questo mezzo per i gameti era impossibile incontrarsi e generare un nuovo individuo, e di fatti possedevano dei sistemi di propulsione che consentivano a loro di “nuotare”. Prova ne è che tutte le piante fin qui elencate, vivevano parzialmente immerse nell’acqua, oppure prosperavano in ambienti decisamente umidi, come del resto fanno le attuali felci e muschi.
Le prime piante dotate di semi erano ancora molto simile alle felci e infatti i paleobotanici le definiscono “felci a seme” o Pteridosperme e risalgono ai periodi del Carbonifero – Giurassico (da circa 199 a 145 milioni di anni fa). Possedevano alcuni caratteri delle felci ed altri tipici delle Gimnosperme (gruppo a cui appartengono le attuali conifere). Quest’ultima classe di piante dominò la flora per quasi tutto il Mesozoico fino alla comparsa delle Angiosperme che restrinse progressivamente la diffusione del loro areale. Al momento attuale si conoscono circa 700 specie di Gimnosperme diffuse soprattutto nelle zone temperate fredde dell’Eurasia e Nordamerica. Sono suddivise in 6 gruppi, alcuni dei quali presenti ancora oggi, altri estintesi nel corso del Mesozoico (Era che iniziò circa 251 milioni di anni fa e si concluse circa 65 milioni di anni fa).
Il gruppo delle Pteridospermofita avevano portamento arboreo con rami che portavano fronde disposte a spirale e si trovavano all’apice di un fusto sottile; i semi erano racchiusi in contenitori a forma di cupola, mentre i tessuti produttrici di polline erano localizzati sulle foglie. Uno dei generi meglio conosciuti tra quelli che erano più ampiamente diffusi nelle foreste carbonifere è Medullosa (fig. 13).
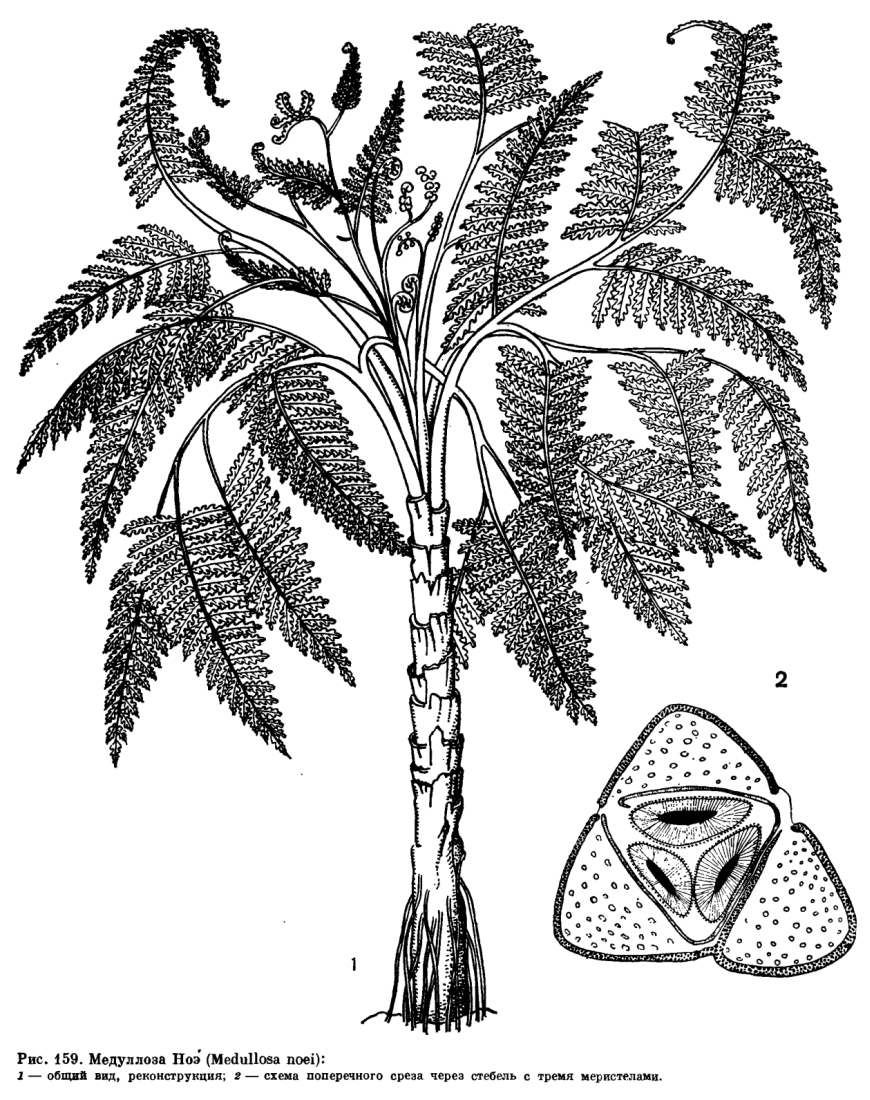
fig. 13
Secondo molti paleobotanici, inoltre, per i loro requisiti morfologici le Pteridosperme si possono ritenere le antenate delle Angiosperme.
Dal gruppo delle Cicadoficali si sono differenziate nel Triassico le Cicadali e le Bennettiali; quest’ultime hanno avuto una grande diffusione nel Mesozoico, però si sono estinte durante il Cretaceo (tra circa 145 e 65 milioni di anni fa). Il loro tronco poteva essere molto corto e rigonfio oppure abbastanza allungato come in Williamsonia (fig. 14).

fig. 14
I semi e i granuli pollinici erano situati all’ascella di foglie modificate disposte a spirale attorno ad un asse principale: possiamo dire la prima pigna (stròbilo) della storia. In alcune specie, addirittura, la base dello stròbilo era avvolta da foglie speciali (brattee), per cui vi erano già tutti gli elementi che compongono un fiore; infatti, la struttura risulta molto simile al fiore della attuale Magnolia, ritenuta la pianta più antica delle Angiosperme, comparsa, infatti, nel Cartaceo inferiore (tra 145 e 99 milioni di anni fa) e giunta sino a nostri tempi (fig. 15).

fig. 15
Un altro gruppo simile al precedente, evolutosi durante il Permiano (periodo che si estende da circa 299 a circa 251 milioni di anni fa), e di cui 10 generi con 100 specie sono attualmente viventi, è quello delle Cycadophyta. Il loro tronco è indiviso come nel genere attuale Cycadales il che conferisce alla pianta un aspetto simile a quello delle palme. Sulla corteccia sono visibili, inoltre, le cicatrici lasciate dalla base delle foglie senescenti che sono cadute; queste, generalmente grandi, sono pennate e riunite in un ciuffo all’estremità apicale del tronco.
Il quarto gruppo, quelle delle Ginkofita (dal Permiano fino ad oggi) raggiunse il suo massimo sviluppo nel corso del giurassico, per poi ridursi progressivamente: oggi è presente solo la specie Ginko biloba (fig. 16) originaria della Cina occidentale. Queste piante sono facilmente riconoscibili per la particolare forma a ventaglio delle loro foglie che presentano ancora un’antica nervatura dicotomica. Inoltre, se si guarda la sagoma della chioma è possibile scorgere il profilo conico che le accomuna con le conifere attuali (fig. 17), ma anche con alcune Angiosperme primitive, quali la già citata Magnolia (fig. 18).
.jpg)
fig. 16

fig. 17

fig. 18
Al gruppo delle Coniferofita (dal Carbonifero fino ad oggi) appartiene la maggior parte delle Gimnosperme viventi; sono diffuse soprattutto nell’emisfero Nord, ad esempio gli abeti, i pini, i larici, i ginepri mancano nell’emisfero Sud. Tuttavia alcuni generi sono propri di tale emisfero, come Podocarpus e Araucaria; in particolare, la specie A. araucana del Cile presenta delle caratteristiche che possono essere ritenute arcaiche come per esempio le foglie spesse triangolari disposte come le tegole di un tetto (fig. 19) e che ricoprono interamente il caule, come abbiamo già visto in precedenza per le briofite più evolute. Si possono distinguere due classi: Cordaitopsida e Coniferopsida. La prima con il genere Cordaites, diffuso ampiamente durante il carbonifero, comprende i più antichi rappresentanti delle conifere, invece, appartengono a Coniferopsida quelle con aspetto piuttosto simile alle specie tuttora viventi. Quest’ultima classe raggiunse il suo massimo sviluppo durante il Giurassico, iniziando via via a declinare nel Cretaceo, in seguito all’evoluzione delle Angiosperme.

fig. 19
Nel Cenozoico (da 65 a 0.3 milioni di anni fa) la crisi si fece sempre più evidente: vi fu una continua restrizione degli areali delle specie e dei generi che in precedenza occupavano territori molto vasti in tutto l’emisfero australe e si ridusse lo sviluppo di nuove forme. Tale declino è legato non solo alle difficoltà che le conifere ebbero nel competere con le angiosperme, ma anche alla ridotta capacità di adattamento ad ambienti diversi del loro patrimonio genetico. Coniferopsida comprende quattro ordini: Lebachiales,Voltzlales, Pinales e Taxales. Le lebachiali (dal Carbonifero al Permiano) erano piante arboree con foglie aghiformi, largamente diffuse nell’emisfero settentrionale. Le voltziali (dal Carbonifero al Giurassico), che mostrano alcune affinità con le conifere viventi, erano forme arboree caratterizzate da foglie squamose, disposte a elica lungo i rami. Le pinali (dal Giurassico ad oggi) sono diffuse in tutto l’emisfero settentrionale. I loro fossili, abbondanti negli strati rocciosi del Cretaceo, sono attribuiti a numerosi generi, tra i quali Pinus, Pseudolarix e Picea. Le taxali (dal Giurassico ad oggi) sono conosciute a partire dal Giurassico grazie ai resti del genere Elatides; Sequoia, tuttora vivente, è noto soprattutto dai fossili rinvenuti in depositi cenozoici; il genere Metasequoia, considerato in pratica un fossile vivente, risale al Cretaceo. Il sesto e ultimo gruppo,quello delle Gnetofita (dal Permiano ad oggi) comprende tre generi molto diversi tra loro; è particolarmente interessante per le caratteristiche anatomiche e fisiologiche che, sotto certi spetti, risultano intermedie tra quelle delle Coniferofita e delle Angiosperme, come per esempio l’impollinazione attraverso gli insetti. Tuttavia, da un punto di vista filogenetico non hanno relazione con gli altri gruppi delle gimnosperme e neppure con quelli delle angiosperme. In effetti, molto interessante è l’unica specie del genere Welwitschia, vive nei deserti dell’Africa orientale e meridionale; è denominata W. Mirabilis (fig. 20) e presenta solo un paio di foglie laminari a crescita continua completamente distese, quasi possedessero una sorta di parentela con le epatiche ancestrali.

fig. 20
Le angiosperme devono il proprio no-me al fatto di possedere semi racchiusi all’interno di un frutto. Estremamente diffuse nella flora attuale, con circa 25000 specie viventi, rappresentano il gruppo più numeroso di piante vascolari. I paleobotanici non sanno ancora con certezza quando siano comparse sulla Terra, anche se è opinione comune che “l’era delle angiosperme” abbia avuto inizio nel Cretaceo superiore, periodo in cui si verificò la loro improvvisa rapida diffusione con forme molto varie, simili ai salici, ai pioppi, agli allori, ai platani e alle magnolie. Nell’arco di pochi milioni di anni, comunque, sostituirono le gimnosperme e le pteridofite come componenti principali delle flore di ogni continente.
Fonti:
Ceruti A. , Il nuovo Pokorny; 1996, Loescher.
I fossili: guida pratica per conoscerli e collezionarli, De Agostini.
Le fotografie n° 17, 18 e 19 sono dell’autore.