Gli ormoni steroidei sono piccole molecole idrofile che, similmente ad altre sostanze attive quali gli ormoni tiroidei e le vitamine A e D, giocano un ruolo molto importante nelle crescita, nella differenziazione cellulare, nella riproduzione e nella morfogenesi degli organismi. La maggior parte di queste azioni sono mediate dal legame di queste molecole a recettori nucleari che agiscono come fattori di trascrizione indotti dal ligando.
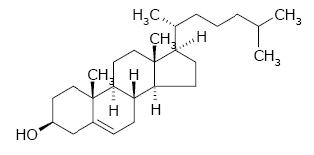
Fig. A. Precursore comune degli ormoni steroidei, il colesterolo. Materiale ottenuto dal Prof. Giorgio Sartor, Università di Bologna a Ravenna
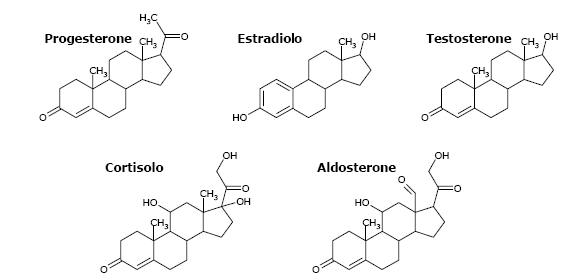
Fig B. Gli ormoni steroidei. Materiale ottenuto dal Prof. Giorgio Sartor, Università di Bologna a Ravenna
I recettori nucleari sono proteine costituite da una serie di domini, chiamati con lettere da A a F (dall'estremità N-terminale alla C-terminale) e contengono tre domini funzionali principali, che operano indipendentemente (vedi Fig C):
- un dominio variabile N-terminale (A e B) che conferisce immunogenicità e modula la trascrizione in maniera specifica al tessuto e al gene attraverso la cosiddetta Activation Function-1 (AF-1);
- Un dominio centrale di legame al DNA (DBD, costituito dal dominio C) che comprende due strutture zinc-finger funzionalmente distinte, attraverso le quali il recettore interagisce fisicamente con l'elica del DNA;
- Il dominio di legame al ligando (LBD, che include il dominio E e, in alcuni recettori, anche l'F), questo contiene la Activation Function-2 (AF-2)
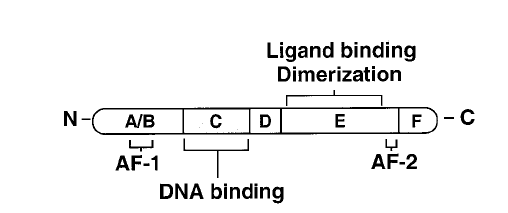
Fig C. Rappresentazione schematica di un recettore nucleare. AF-1, dominio di attivazione indipendente dal ligando; DBD, dominio id legame al DNA; AF-2, dominio di attivazione dipendente dal ligando. Tratto da ARANDA & PASCUAL, PHYSIOL. REV. Vol. 81, No. 3, July 2001
Gli ormoni steroidei agiscono attraverso la regolazione della trascrizione del DNA in RNA messaggero, processo che può avvenire solo a seguito dell'attivazione di alcuni elementi chiave presenti nella regione regolatoria del gene. Questi elementi sono collocati sullo stesso tratto di DNA che contiene il gene che regolano e sono suddivisi in 4 gruppi principali: promotori, enhancer sensibili all'ormone, silenziatori e enhancer insensibili agli ormoni.
Il promotore è localizzato vicino al sito di inizio della trascrizione ed è costituito da due elementi: il TATA box e il promotore a monte.
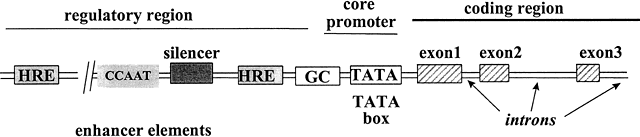
Fig. D. Schema della regione regolatoria e strutturale di un gene tipo regolato dagli ormoni steroidei. Adattato da Tsai M-J, O'Malley BW. Molecular Mechanism of action of steroid/throid receptor superfamily membres. Annu Rev Biochem 1994; 63:451-483.
I recettori nucleari regolano la trascrizione legandosi a specifiche sequenze di DNA presenti all'interno dei geni bersaglio, sequenze che vengono chiamate elementi di risposta all'ormone (HRE). Gli HRE presentano un motivo centrale di 6 paia di basi per il riconoscimento del recettore. Sebbene alcuni recettori possano legarsi come monomeri a un singolo motivo esamerico, la maggior parte si legano come omo- o etero-dimeri a una coppia di motivi esamerici (palindrome, palindrome inverse o ripetizioni dirette). Due recettori monomerici si legano in maniera cooperativa ai relativi elementi di risposta e le interfacce di dimerizzazione sembrano essere collocate sia nel LBD che nel DBD.
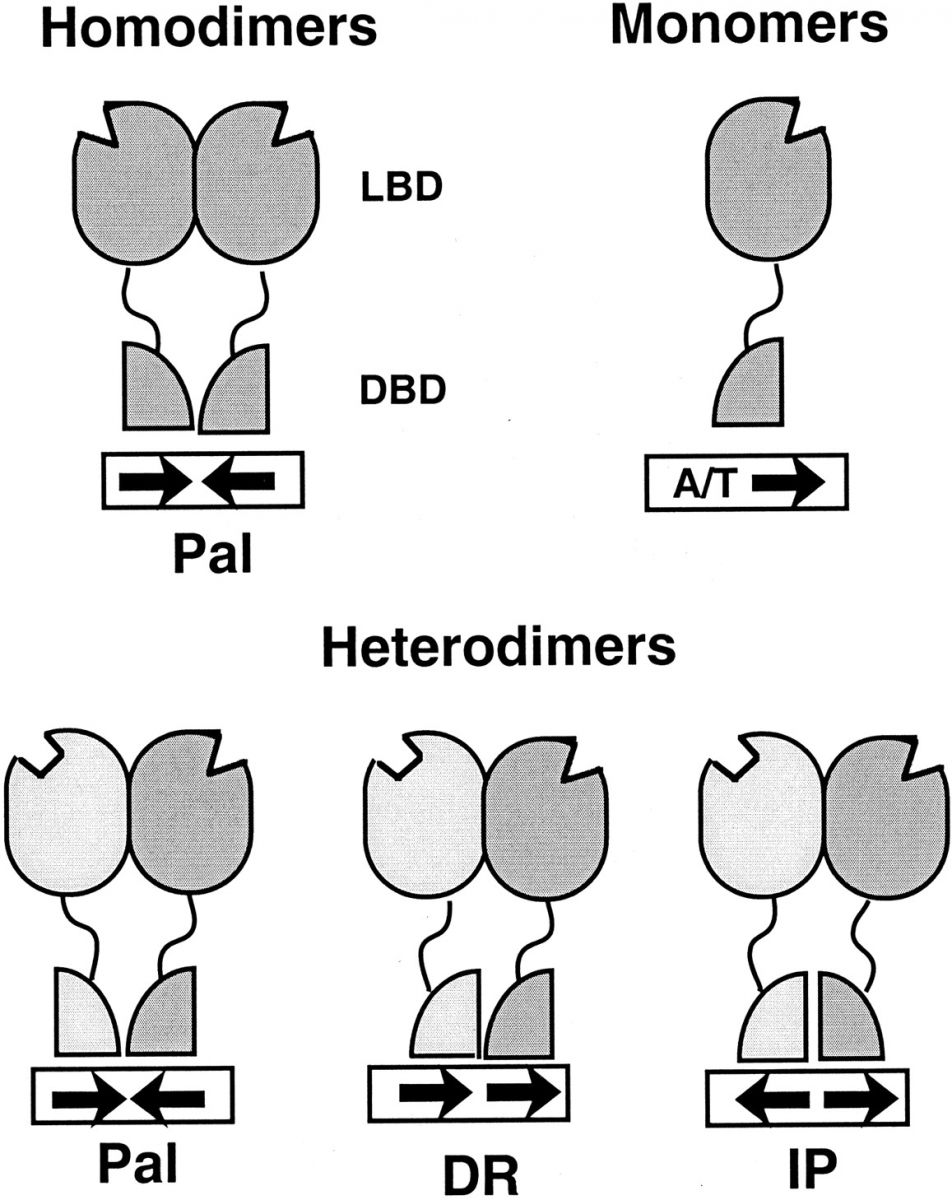
Fig E. Schema di legame del recettore agli HRE. I recettori possono legare il DNA come monomeri, omodimeri o eterodimeri. La dimerizzazione è mediata da ripetizioni di 7 aminoacidi fortemente idrofobici presenti nel LBD e il legame cooperativo dei dimeri recettoriali è facilitato dalla struttura DNA-dipendente che si forma dall'unione dei due DBD. I recettori steroidei si legano come omodimeri in maniera simmetrica a elementi palindromi, spaziati da tre nucleotidi. Il legame monomerico richiede sul DNA una semi-sequenza base preceduta da una regione ricca in A/T. Gli eterodimeri possono riconoscere differenti HRE, nei quali la sequenza base può essere organizzata in palindromi (Pal), ripetizioni dirette (DRs) o palindromi invertiti (IPs). Tratto da ARANDA & PASCUAL, PHYSIOL. REV. Vol. 81, No. 3, July 2001.
La principale funzione del dominio di legame al DNA (DBD) è di legarsi a specifici elementi di risposta all'ormone (HRE) presenti sul gene bersaglio. Il legame al DNA avviene attraverso la coordinazione tetraedrica dello zinco di 4 cisteine presenti in ciascuna delle due estensioni che formano due zinc finger strutturalmente distinti. La specificità di legame del HRE è determinata dal primo zinc finger, che è il più conservato, mentre il secondo zinc finger è coinvolto nella dimerizzazione e nella stabilizzazione del legame al DNA attraverso le interazioni ioniche con lo scheletro fosfato del DNA.
Nel dettaglio, l'azione degli ormoni steroidei avviene in 6 stadi:
- Ingresso dell'ormone nelle cellule del tessuto bersaglio
- Localizzazione intracellulare dell'ormone e del recettore nucleare
- Ingresso del recettore nucleare legato all'ormone nel nucleo
- Attivazione:
- ligando dipendente del complesso recettore-ormone
- ligando indipendente del recettore nucleare
- Legame al DNA
- Regolazione della trascrizione genica:
- regolazione diretta della trascrizione
- regolazione indiretta attraverso l'interazione del complesso con altri fattori di trascrizione
- repressione della trascrizione genica
1- ingresso dell'ormone nelle cellule del tessuto bersaglio
Gli ormoni steoridei sono liposolubili e circolano nel flusso sanguigno liberi o legati a proteine plasmatiche. Le proteine di legame extracellulare possono anche fungere da modulatori della risposta ormonale, limitando la quantità di ormoni disponibili per la cellula.
Quindi gli ormoni steroidei possono entrare legati a globuline attraverso endocitosi mediata da recettore oppure diffondere liberamente nel citoplasma.
2- localizzazione intracellulare dell'ormone e del recettore nucleare
La maggior parte dei recettori, quando non impegnati nel legame con l'ormone, risiedono nel nucleo. Fanno eccezione i recettori per i glucocorticoidi e i mineralcorticoidi, che si trovano nel citoplasma in associazione con varie proteine, tra le quali le chaperonine hsp70 e hsp90. A seguito dell'attivazione dovuta al legame con l'ormone e il rilascio delle proteine associate, il complesso monomerico ormone/recettore dimerizza e trasloca nel nucleo.
3- Ingresso del recettore nucleare legato all'ormone nel nucleo
I recettori degli ormoni entrano nel nucleo a seguito di due processi: diffusione passiva attraverso il canale centrale dei pori nucleari o trasporto attivo mediato da recettore.
4- Attivazione:
a. ligando-dipendente del complesso recettore-ormone
Il legame del recettore con l'ormone porta alla dissociazione delle proteine associate e alla fosforilazione del recettore, evento che potrebbe giocare un ruolo molto importante nel legame del recettore al proprio specifico elemento di risposta localizzato sul gene bersaglio.
b. ligando indipendente del recettore nucleare
La fosforilazione può avvenire anche in assenza del ligando, a seguito di cascate di segnale derivanti dalla membrana cellulare.
5- legame al DNA
Quando il complesso ormone/recettore si lega al HRE di competenza, deforma il DNA, causandone una curvatura che probabilmente facilita l'interazione tra le componenti del complesso di trascrizione che si legano a diversi siti e promuove la creazione di anse nel DNA per permettere alle singole proteine di entrare in contatto con molteplici elementi del DNA.
6- Regolazione della trascrizione genica:
a. regolazione diretta della trascrizione
I recettori degli ormoni steroidei interagiscono con i fattori di trascrizione base attraverso le regioni AF-1 e AF-2.
Inoltre esistono molti differenti coattivatori che interagiscono direttamente con il complesso recettore/ormone e sostengono la trascrizione.
Uno dei modelli attualmente considerati validi, suggerisce che le differenti cellule bersaglio esprimano diversi livelli di coattivatori e corepessori che, insieme con la quantità di proteine recettoriali e di ligando, permettono una fine regolazione della trascrizione genica in risposta agli ormoni steroidei.
b. regolazione indiretta attraverso l'interazione del complesso con altri fattori di trascrizione
I recettori degli ormoni steroidei possono interagire direttamente con diversi fattori di trascrizione e alterare così la trascrizione genica, senza che il recettore interagisca direttamente con il DNA.
c. repressione della trascrizione genica
Il legame del complesso recettore/ormone può anche reprimere la trascrizione di alcuni geni bersaglio, anche se il meccanismo non è stato ancora chiarito completamente.
Una possibilità è che il recettore per lo steroide si leghi sul DNA allo stesso sito di legame utilizzato dalle proteine attivatrici, bloccandone completamente l'accesso.
Alternativamente, la repressione dei recettori steroidei può avvenire attraverso il legame al HRE e la collezione di proteine copressori che limitino l'attività degli attivatori legati ai promotori.
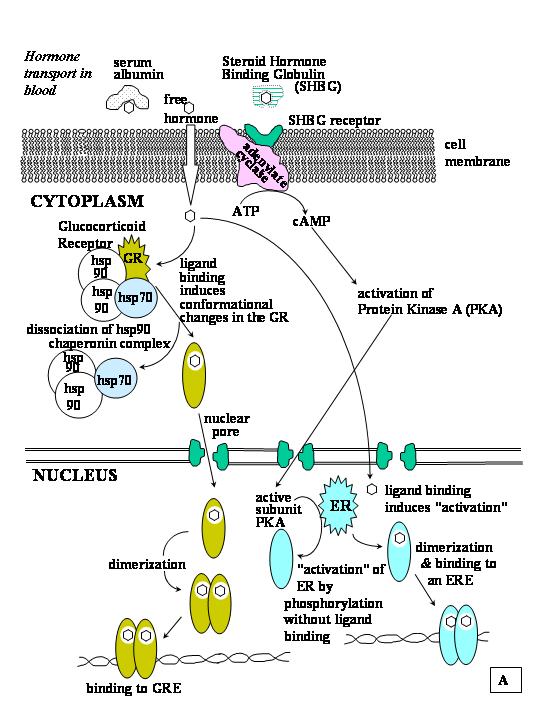
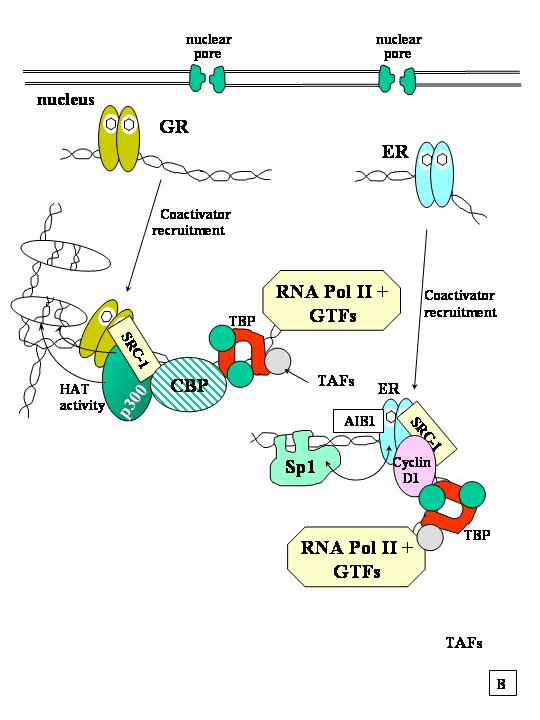
Fig F. Meccanismi di azione dei recettori per gli ormoni steroidei nelle cellule bersaglio. A. molti ormoni steroidei che entrano nella cellula viaggiano nel flusso sanguigno o liberi o blandamente legati all'albumina serica. Gli estrogeni e gli androgeni invece si legano con alta affinità a globuline specifiche (SHBG). La membrana cellulare contiene un recettore per SHBG associato a una adenilato ciclasi. L'attivazione della adenilato ciclasi genera cAMP che attiva la protein chinasi A (pKA) e la conseguente cascata di fosforilazione. B.Una volta legato all'HRE, il recettore induce una deformazione del DNA e richiama proteine attivatrici. Klinge, C., Rao, C., Glob. libr. women's med., (ISSN:1756-2228) 2008.
Riferimenti e bibliografia:
Klinge, C., Rao, C., Glob. libr. women's med., (ISSN:1756-2228) 2008.
C. Cooper et al.,Biochimie 2011 doi: 10.1016/j.biochi.2011.07.002
Gronemeyer H., 1992, The FASEB Journal
John W. Honour, J Clin Res Ped Endo 2009;1(5):209-226. DOI: 10.4274/jcrpe.v1i5.209
http://www.pnas.org/content/suppl/2007/12/18/0710487105.DC1
http://physrev.physiology.org/content/81/3/1269.full-text.pdf+html