ll tema è enorme ed è difficile sapere fino a che punto di approfondimento ci si deve spingere. La trattazione sarà quindi abbastanza generale, spero che sia sufficiente. I link sono, purtroppo, tutti in inglese.
Parliamo di infezioni e di risposte immunitarie dell’organismo, facendo un breve riepilogo.
http://www-immuno.path.cam.ac.uk/~immuno/part1/lec15/lec15_97.html
Esistono dei sistemi di risposta immediati:
- il primo sistema di difesa è sotto forma di molecole preformate, già presenti in modo costitutivo. Il difensore immediato più importante è il C3. Il C3 viene generato in maniera continua ed è un componente del complemento. ll complemento da solo riesce a distrugge alcuni batteri, soprattutto Gram+.
- Un secondo sistema è quello degli anticorpi naturali già presetni nell’organismo, cioè anticorpi aspecifici, in grado comunque di bloccare alcuni agenti infettanti.
- Una terza difesa è rappresentata dai neutrofili nel sangue e dai macrofagi, che si trovano in tutti i tessuti, soprattutto mucose. Ambedue hanno dei recettori che permettono loro di legare e fagocitare i microbi. Questi recettori riconoscono certe strutture come i lipopolisaccaridi dei Gram –
Questo “esercito ” di armi già pronte non è sufficiente per cui c’è una prima risposta immunitaria, che si manifesta a partire da 4 ore dall’infezione e prevede la sintesi di composti ex-novo.
- I macrofagi, che iniziano a produrre citochine. In particolare producono TNalfa, IL2 e IL1.
- I NK – natural killer – sono linfociti attivati dalle citochine. In genere attivi contro i virus.
- Gli interferoni, che limitano la infezioni virale nelle fasi precoci, in attesa che si formino gli anticorpi specifci.
Dopo quattro giorni vengono formati gli anticorpi specifici e i linfociti T.
Riprendendo il discorso sui Gram negativi, iniziamo dalla composizione della parete in cui è inglobata la tossina e dalla struttura della tossina, che è un lipopolisaccaride o LPS.
http://www.bact.wisc.edu/Microtextbook/index.php?module=Book&func=displayarticle&art_id=60
Da: http://www.bmanuel.org/courses/membr+_.jpg
{kind=link}
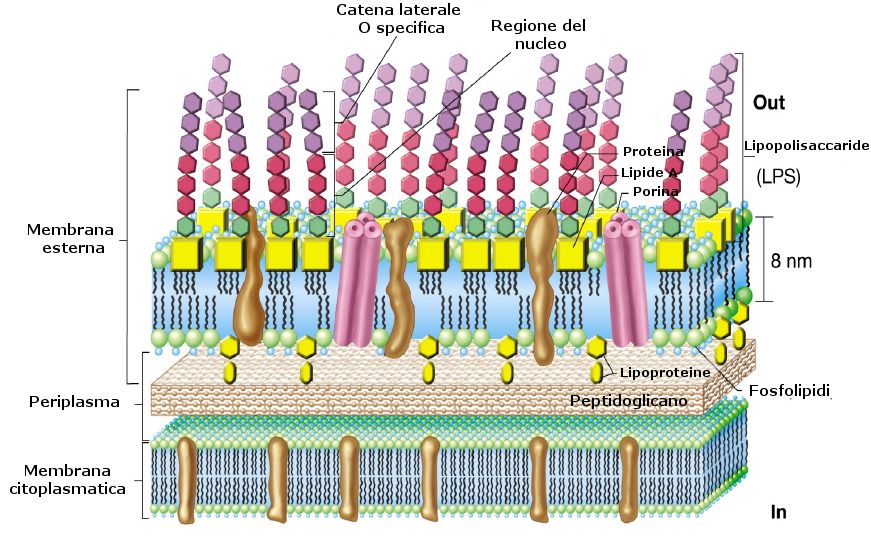
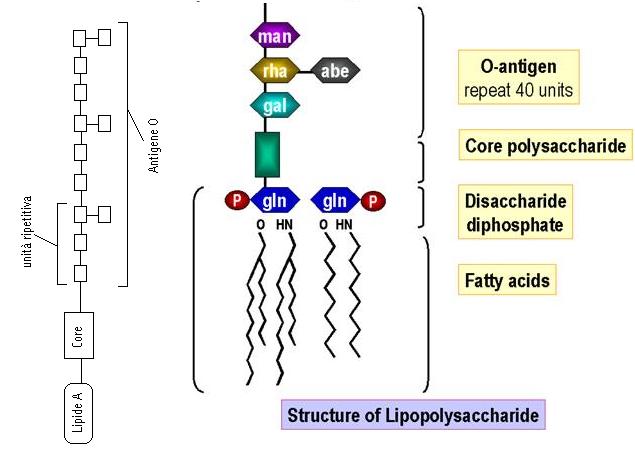
Il lipide A è costituito da una molecola di Glucosamina esterificata e fosforilata e costituisce la tossina vera e propria. Il core è invece costituito dall’acido chetodeossioctonico e da un eptoso. Il core mantiene invariata la sua struttura in tutti i batteri.
L’antigene O invece è costituito da una lunga catena polisaccaridica e varia in ogni batterio.
Il meccanismo d’azione della tossina consiste nella attivazione dei macrofagi che a loro volta produrrano dei mediatori.
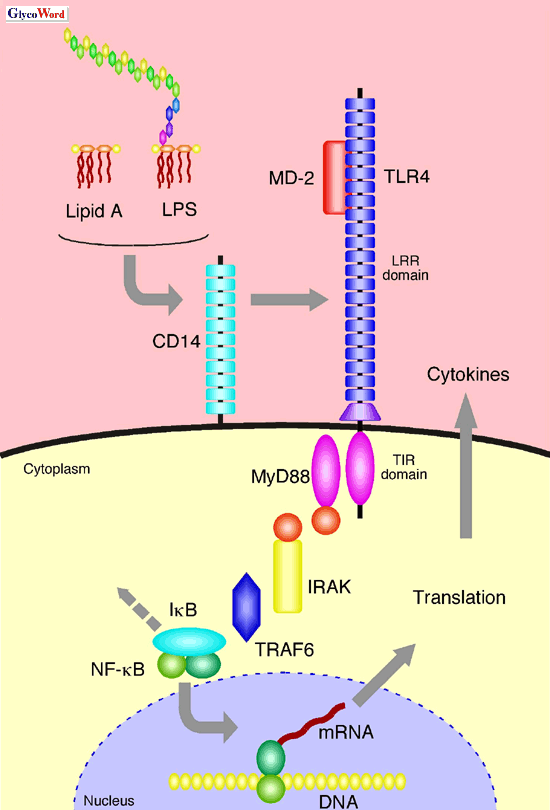
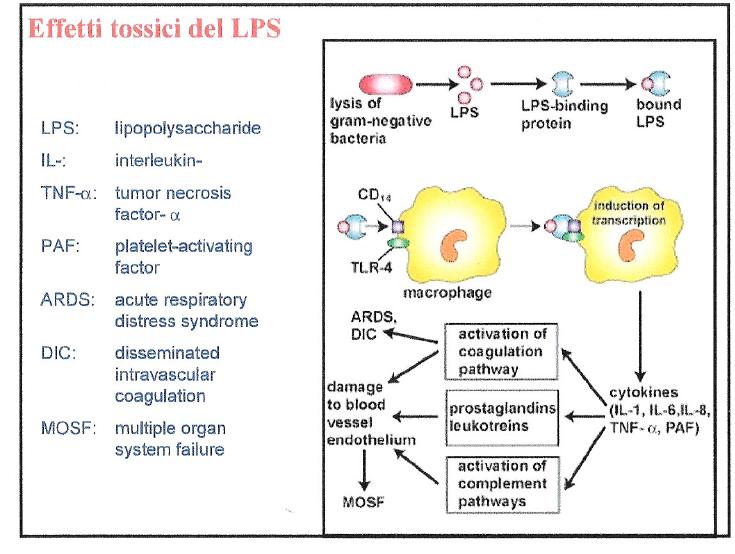
Secondo quanto riportato inquesto articolo:
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&list_uids=15949139&dopt=Abstract
La endotossina dei Gram- è inserita nella parete cellulare del micororganismo e bisogna quindi che venga liberata. La proteina LBP e CD14 si combinano, per estrarre singole molecole di endotossine dalla parete del batterio. Si forma un complesso che serve a trasferire il lipide A alla proteina MD -2 . Il complesso MD 2 – LPS attiva la proteina TLR4 a concentrazioni picomolari.
TLR4 fa parte di una serie di proteine transmembrana o TLRs, con un dominio esterno ricco di residuidi leucina o LRR e di un dominio interno alla cellula detto TIR, che trasduce (passa) il segnale.
Il segnale passa alla proteina MyD88, che a sua volta tramite la chinasi IRAK, stimola la attività di ligasi della TRAF6.
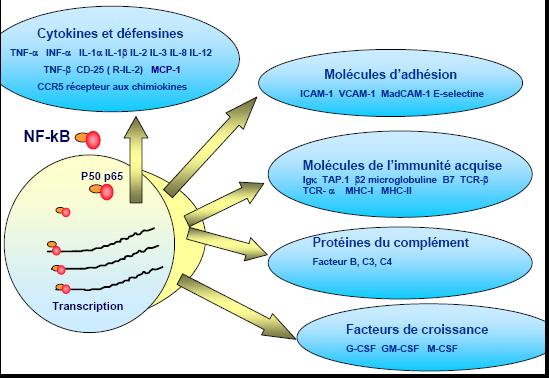
Questa a sua volta attiva dei fattori di trascrizione NF-kB che controllano la trascrizione. Il Nuclear Factor- kB (Nf-k B) è un fattore di trascrizione (FT) ubiquitario conservato nelle cellule eucariotiche. Nf-k B è attivato da numerosi stimoli compresi prodotti virali e batterici, radiazioni ultraviolette, radicali ossidanti, citochine, sostanze chimiche varie. Una volta attivato, Nf-k B controlla direttamente, o con la cooperazione di altri fattori di trascrizione, l’attività di oltre 100 geni che generano citochine, fattori di crescita, chemochine, molecole di adesione, proteine della fase acuta.
La prolungata attivazione di NFkB può causare gravi condizioni infiammatorie e persino la morte da eccessiva produzione di citochine; pertanto è essenziale non solo che NFkB sia rapidamente attivato nel corso delle infezioni, ma anche che la sua attivazione abbia rapidamente termine subito dopo che lo stimolo infiammatorio/infettivo sia stato allontanato.
All’interno delle cellule vi sono, pertanto, meccanismi di controllo che ne limitano l’attivazione.