La polarità è stata recentemente definita in contesto biologico come una distribuzione persistente e asimmetrica di strutture lungo un asse (Cove et al., 1999) e questa definizione è applicabile alla polarità a livello di organelli, di cellule, di organi e di interi organismi. L’asse di allineamento non specifica la direzione della polarità ma solo la relazione tra gli assi e alcuni punti di riferimento esterni. La direzione in cui la distribuzione asimmetrica e ordinata delle strutture si presenta, viene definita come l’orientamento dell’asse. Quindi due oggetti polari potrebbero essere entrambi orientati secondo un gradiente esterno, ad esempio la forza di gravità, ma orientati in direzioni opposte. Lo stabilirsi e il mantenimento della polarità cellulare si basa su una distribuzione asimmetrica del citoscheletro e dell’attività di altre proteine all’interno della cellula. La concentrazione di specifiche proteine in una determinata regione cellulare è parte essenziale del processo. Le proteine associate alla membrana e quelle secrete sono indirizzate durante la traduzione nel reticolo endoplasmatico, quindi traslocano nell’appropriato compartimento attraverso l’apparato di Golgi e le vescicole che da esso si staccano. Al contrario, molte proteine citoplasmatiche sono posizionate attraverso la pre-localizzazione dei loro trascritti. Dal momento che un singolo mRNA è stampo per più copie della proteina, localizzare i trascritti è un meccanismo efficiente per generare una fonte localizzata e concentrata di proteine. Quando accoppiato con la localizzazione del mRNA, il controllo localizzato della traduzione può portare a un controllo spaziale e temporale dell’espressione genica all’interno della cellula. Inoltre, la restrizione locale e temporale dell’attività proteica tramite la traduzione sito-dipendente può servire per prevenire la comparsa di interazioni potenzialmente pericolose in altri comparti della cellula.

Esempio di cellula polarizzata: il neurone. Colorazione:impregnazione argentica.
Gli esempi di cellule polari sono molteplici in tutti i Metazoi: le cellule epiteliali e le cellule embrionali sono gli elementi polari pù� ampiamente studiati.
Le cellule epiteliali sono polarizzate lungo il loro asse apico-basale e i loro compartimenti di membrana apicale e basolaterale si separano durante lo sviluppo. La membrana cellulare delle cellule epiteliali mature è divisa in due domini: un dominio apicale che si affaccia sull’ambiente esterno e un dominio basolaterale che contatta l’ambiente interno e la zona vascolarizzata. All’interno dei tessuti epiteliali maturi, la polarità cellulare regola la morfologia cellulare, i segnali intracellulari, la divisione cellulare asimmetrica, la migrazione cellulare, la fisiologia cellulare e istologica così come la complessa organogenesi. Molti epiteli sono polarizzati anche lungo il loro asse ortogonale all’asse apico-basale, portando a una asimmetria all’interno del piano dell’epitelio (polarità cellulare planare, PCP, o polarità tissutale). La polarizzazione lungo questo asse richiede un segnale per raggiungere ampi volumi di cellule. Il segnale genera una asimmetria molecolare all’interno delle cellule bersaglio e quindi converte questa asimmetria in una riorganizzazione citoscheletrica.

Esempio di tessuto epiteliale pseudstratificato ciliato
La maggior parte delle proteine di membrana nelle cellule epiteliali sono distribuite principalmente o sul dominio di membrana apicale o su quello basolaterale, ma poche sono distribuite su entrambe le superfici. La polarità della membrana nelle cellule epiteliali è stata studiata utilizzando glicoproteine del capside virale come marcatori. Nelle cellule epiteliali, i virus dell’influenza indirizzano le loro glicoproteine verso la superficie apicale della membrana che si affaccia sul lume, identificandola. Nell’epitelio i differenti domini della membrana plasmatici sono chiaramente separati da giunzioni strette in grado di mantenere la polarità della membrana.
L’analisi in Drosophila delle mutazioni di DSH che distruggono la via di segnalazione della PCP hanno dimostrato che esiste un dominio specifico che agisce a livello della PCP. La via di segnalazione di PCP comprende un certo numero di componenti principali, i cui ruoli vengono suddivisi durante la polarizzazione dei diversi tessuti bersaglio, pi�ùalcuni componenti addizionali specifici i cui ruoli sono più specializzati.
La distribuzione asimmetrica delle proteine responsabili della determinazione e del mantenimento della polarità sono stati studiati particolarmente in dettaglio nel moscerino della frutta Drosophila, che presenta numerose strutture polarizzate, quali le ali, le appendici e gli occhi composti.
Lavori recenti hanno dimostrato che un complesso di segnalazione di Fz viene assemblato simmetricamente attorno alle cellule e in seguito diviene asimmetricamente localizzato durante la generazione della polarità nelle ali pupali di Drosophila. Sia Fz che Dsh sono stati dimostrati localizzare al confine distale delle cellule delle ali pupali, mentre Flamingo (Fmi), conosciuto anche come StarryNight (Stan), un membro della super famiglia delle caderine, si pensa sia localizzato ai confini prossimale e distale. Diego (Dgo), una proteina con ripetizioni di ankirina, si localizza in questi confini. Un controllo a feedback sembra mediare l’acquisizione di questa asimmetria. Le mutazioni che la localizzazione asimmetrica di questi componenti produce anche polarità planari aberranti. Questa correlazione suggerisce che la loro localizzazione subcellulare asimmetrica serva come segnale per la direzione della riorganizzazione citoscheletrica.
Un modello proposto per spiegare l’insorgenza della polarità, prevede che un simile segnale asimmetrico giochi un ruolo nel generare la polarià delle cellule precursori delle setole e degli ommatidi. Il segnale asimmetrico sarebbe legato, attraverso l’azione di fattori tessuto-specifici, a elementi che controllano i cambiamenti morfogenetici peculiari che si osservano in ciascun tessuto. Mentre la localizzazione subcellulare di queste proteine non è stata riportata nell’occhio, fmi/Stan non mostra localizzazione asimmetrica nelle cellule precursori delle setole che sono il bersaglio della via di segnalazione di fz. Inoltre, mutazioni in Dgo non sembrano influire la polarità delle setole. Quindi, non è chiaro se una simile localizzazione asimmetrica di queste molecole segnale giochino un ruolo nel determinare la polarità di tutti i tessuti.
La polarità è centrale per lo sviluppo delle piante e degli animali, sia nella loro evoluzione che per la produzione della complessità individuale. La polarità a livello della singola cellula è al centro dello sviluppo degli organismi pluricellulari complessi, dal momento che la divisione di una cellula polare genera cellule figlie non equivalenti, permettendo il successivo sviluppo differenziato.
Sebbene in molti sistemi, la generazione di un asse polare richieda un input asimmetrico, alcune strutture biologiche polari si sviluppano in maniera autonoma. I capsidi asimmetrici dei virus, come quelli del ceppo T, possiedono una struttura estremamente polare che è ottenuta dall’assemblamento dei componenti del capside senza la necessità di un input polare esterno. Anche gli organelli, quali i ribosomi, possono raggiungere la polarità senza un input esterno. Un ulteriore esempio è dato dal differenziamento di una cellula figlia in una pre-spora e l’altra in una cellula sporigena, all’inizio della sporulazione nel batterio Bacillus subtilis, asimmetria che si pensa sia raggiunta indipendentemente da input esterni. La polarità del microgametofito (granulo di polline) delle piante da fiore è correlato all’asse prossimo-distale delle microspore da cui esso si sviluppa. Sebbene questa relazione possa variare tra le specie, per una determinata specie essa si mantiene costante, indicando che la polarità del granulo pollineo deve la sua origine alla topologia di formazione della tetrade di microspore.
Lo sviluppo dell’asse polare tuttavia richiede, nella maggior parte dei casi, un input prodotto da un segnale biologico o dall’ambiente fisico, in particolare un lieve gradiente o uno stimolo gravitario. In molti sistemi animali, incluso il nematode Caenorhabditis elegans, e gli anfibi come Rana, il punto di entrata dello spermatozoo nell’uovo fornisce lo stimolo che rompe la simmetria. Il punto di entrata dello spermatozoo è anche lo stimolo base per la determinazione dell’asse polare nello zigote in sviluppo dell’alga fucoide Pelvetia compressa, tuttavia, se lo zigote sviluppa in presenza di un gradiente di luce, l’asse può essere ri-specificato con un allineamento differente. Sembra che il punto di ingresso dello spermatozoo sia in molti sistemi casuale, quindi un segnale biologico casuale è responsabile dell’inizio del programma che porta alla stabilirsi dell’asse polare. La determinazione dell’allineamento dell’asse dorso-ventrale nell’embrione del moscerino della frutta, è un altro esempio in cui sembra che un evento biologico casuale generi la polarità. Precocemente nella maturazione, il nucleo dell’oocita migra dalla sua posizione originaria mediale a una posizione anteriore. L�asimmetria che porta allo stabilirsi dell’asse dorso-ventrale è assicurata dalla posizione casuale occupata dal nucleo dell’oocita dopo la migrazione. La trascrizione nel nucleo dell�oocita del gene gurken porta alla produzione localizzata di una molecola secreta che segnala alle cellule materne ad esso vicine, mettendo in moto il meccanismo che specifica il lato dorsale dell’embrione. L’input biologico può, tuttavia, essere più difficile da puntualizzare, lasciando ignota l’origine dell’asimmetria. Anche per il sistema animale più studiato, Drosophila melanogaster, mancano ancora idettagli su come venga generato l’asse antero-posteriore dell’embrione. La determinazione degli assi coinvolge l’organizzazione di una distribuzione asimmetrica degli mRNA nell’oocita. Questi mRNA sono trascritti nei nuclei del tessuto materno che circonda l’oocita e sono trasportati nell’oocita dove vengono localizzati. Il prodotto del gene nannos, per esempio, si localizza all’estremità dell’oocita destinata a divenire la porzione posteriore dell’animale. Alcune prove indicano che il trasporto polare richiesto dipende da microtubuli polarizzati pre-esistenti. Se così fosse, deve essere a sua volta spiegato il meccanismo responsabile della determinazione della polarità del citoscheletro. Probabilmente, alla fine, il programma per determinare l’asse antero-posteriore deve la sua origine all’anatomia materna.

L’oogenesi in Xenopus laevis (da http://www.xenbase.org/cell/cyto/oocyte.htm)
Sebbene la polarità sia universale negli organismi pluricellulari, dal momento che porta a funzioni cellulari differenziate e alla formazione degli organi, i requisiti per la generazione della polarità dipenderanno dal fatto che l’organismo in sviluppo sia mobile o no. Un organismo mobile è in grado di aggiustare il suo orientamento nel mondo fisico. Negli organismi mobili, un asse polare ha un valore adattativo, con organi per la percezione sensoriale e per l’analisi dell’ambiente concentrati nella zona del corpo che incontra l’ambiente per prima, convenzionalmente quella anteriore. Per gli organismi mobili terrestri, gli organi locomotori sono ventrali, in una posizione in cui possono interagire con il suolo. Tuttavia non c’è necessità per un organismo mobile di svilupparsi orientato con la forza di gravità dal momento che può aggiustare il suo orientamento in seguito. Quindi, sebbene l�allineamento dell’asse dorso-ventrale in Drosophila sembri basato sul caso, è solo necessario organizzare questo asse ad angolo retto rispetto all’asse antero-posteriore, essendo l’allineamento sul piano perpendicolare non importante.
Seme e embrione( http://wappingersschools.org/)
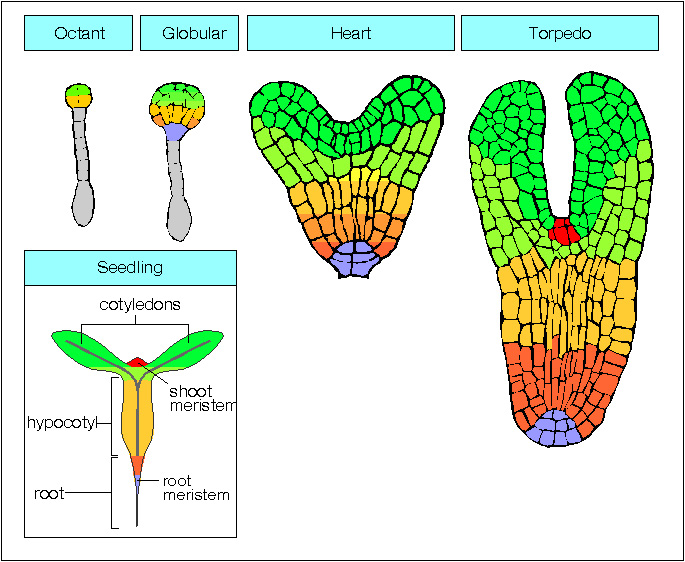
Sviluppo di una pianta (da http://www.mun.ca/biology/desmid/brian/BIOL3530/DB_Ch07/DBNPlant.html)
Le piante, essendo non mobili, possiedono necessità maggiori per gli input ambientali, ma in molti casi la necessità non riguarda l’allineamento e l’orientamento dell’asse polare con l’ambiente fisico nel momento della sua determinazione, ma è modificabile in risposta agli input ambientali. Quindi l’embrione all’interno del seme non richiederà un allineamento dei suoi assi polari in base alla gravità o alla luce nel momento in cui questi vengono determinati, ma è indispensabile che possa rispondere a questi input durante la germinazione. Solo dove non avverranno altri movimenti dopo lo sviluppo polare, sarà utile la presenza di un controllo diretto dell’ambiente esterno durante la generazione degli assi e anche qui gli assi potrebbero essere stabiliti inizialmente senza input ambientali e successivamente modificati nell’allineamento in risposta alla luce o alla gravità.
Sito sui geni implicati nella polarità planare link
Sito polarità cellulare link