I linfociti B e T sono le uniche cellule del sistema immunitario dotate di recettori specifici per gli antigeni, che sono prodotti dei diversi microbi o molecole di natura non infettiva attraverso cui sono riconosciuti. I linfociti T riconoscono solo frammenti di antigeni proteici esposti da cellule specificamente deputate a presentare l’antigene, dette APC. Si suddividono in linfociti T helper, che tra le varie funzioni aiutano i linfociti B a produrre anticorpi, e linfociti T citotossici o CTL, che invece uccidono cellule che albergano microbi intracellulari. I linfociti B esprimono sulla superficie e producono molecole di anticorpo, specifiche proteine da loro prodotte; non hanno bisogno di presentazione per riconoscere gli antigeni, che possono essere proteine, polisaccaridi, acidi nucleici o lipidi.
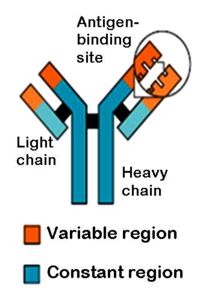
Gli anticorpi si trovano in forma di recettore di membrana (chiamato BCR) quando svolgono il compito di riconoscere l’antigene, oppure in forma solubile (immunoglobuline o Ig) nella fase effettrice. Sono formati da due catene pesanti e due leggere, identiche a due a due: le catene pesanti sono costituite da tre o quattro domini costanti C e uno variabile V, quelle leggere da uno e uno; esistono due tipi di catena leggera (κ e λ) e cinque di catena pesante (α, γ, δ, ε, μ), diverse a livello del dominio costante: in base al tipo di catena pesante si definiscono cinque classi o isotipi di Ig (A, C, D, E, M). La regione costante è comune a tutte le Ig appartenenti allo stesso isotipo ed ha il significato di biologico di portare a risposte immunitarie più efficaci secondo la natura degli agenti patogeni (virus, batteri, protozoi); quella variabile è il sito di combinazione con l’antigene, perciò conferisce specificità per il particolare patogeno riconosciuto. Poco conosciute sono le risposte immunitarie causate da antigeni non proteici (risposte T-indipendenti); si sa invece che nel caso antigeni proteici la specificità recettoriale e la determinazione della classe si generano nel corso di un processo di attivazione linfocitaria mediato da linfociti T helper (risposte T-dipendenti).
Da: http://it.wikipedia.org/wiki/Anticorpo
Tutti i linfociti si generano nel midollo osseo, ma i linfociti T maturano nel timo (organi linfoidi primari). Gli organi linfoidi periferici sono invece milza, linfonodi ed il sistema immunitario associato a cute e mucose,cioè i siti dove i linfociti risiedono prima di incontrare l’antigene; a questo stadio sono detti vergini, in quanto non sono completamente differenziati e sono ancora incapaci di scatenare la risposta immunitaria. L’attivazione è frutto di una cooperazione tra linfociti T helper e linfociti B: al centro dei linfonodi, i primi riconoscono l’antigene presentato da APC, proliferano, si differenziano e migrano verso la periferia; i secondi a loro volta riconoscono un antigene, che può essere differente, ma ugualmente specifico per lo stesso agente patogeno, lo processano e migrano verso il centro del linfonodo. L’interazione avviene in quanto i linfociti B si comportano come APC nei confronti dei linfociti T, presentando l’antigene, mentre questi ultimi esprimono sulla superficie un ligando (CD40L) specifico per un recettore dei linfociti B (CD40). Ciò induce la proliferazione e il differenziamento del linfocita B in cellula che produce anticorpi o plasmacellula. Tale processo è amplificato dalla contemporanea produzione, da parte dei linfociti T, di citochine, peptici usati nella comunicazione cellulare: in questo caso diverse citochine condizionano lo scambio verso determinate classi e la maturazione dell’affinità linfocitaria.
Per quanto riguarda lo scambio di classe, in assenza di stimolazione i linfociti B vergini esprimono IgD, che sono prodotte solo a questo stadio, e IgM, entrambe come recettori di membrana. Le varie citochine promuovono diverse ricombinazioni del locus genico del dominio costante che portano alla perdita di tutti i geni delle catene pesanti meno uno, che è poi riarrangiato con il gene del dominio variabile e genera la catena pesante, determinando la classe; lo stesso avviene per la catena leggera. Per quanto riguarda la specificità, esiste un processo chiamato maturazione dell’affinità attraverso cui l’affinità degli anticorpi aumenta al prolungarsi o al ripetersi dell’esposizione all’antigene. Nei linfociti in proliferazione si verifica un aumento del tasso di mutazione a carico delle regioni V degli anticorpi prodotti, in particolare a livello delle regioni ipervariabili (CDR) deputate al legame con l’antigene: questo fenomeno è chiamato ipermutazione somatica e porta alla generazione di numerosi cloni le cui Ig hanno affinità variabile all’antigene. Parallelamente, i primi anticorpi secreti si legano all’antigene e creano dei complessi catturati dalle cellule follicolari dendritiche (FDC, ), che processano ed espongono l’antigene: se i cloni dei linfociti B non riconoscono l’antigene vanno incontro ad apoptosi. Via via che la risposta si sviluppa, o a seguito di immunizzazioni ripetute, la quantità di anticorpi prodotti aumenta, mentre quella di antigene diminuisce di conseguenza: i linfociti selezionati devono quindi legare l’antigene con affinità crescente. Buona parte di questi cloni svolge subito il proprio compito, ma un piccolo numero si comporta da cellule della memoria che restano in circolo per lungo tempo, in modo tale che se si ripresenta lo stesso agente patogeno l’organismo ha già sviluppato delle cellule pronte a scatenare nuovamente la risposta immunitaria, che è più veloce ed efficace rispetto alla prima volta.
Per quanto riguarda i meccanismi di eliminazione, innanzitutto gli anticorpi neutralizzano i microbi legandosi alle strutture che usano per penetrare nell’ospite, poi li rivestono (un processo detto opsonizzazione) ed infine ne facilitano la fagocitosi in cellule specializzate (macrofagi e neutrofili); queste cellule poi possono essere indotte da linfociti T helper a degradare i patogeni oppure uccise dai linfociti T CTL. In alternativa, altre cellule con attività citotossica come le cellule NK possono legarsi a cellule rivestite da anticorpi e causarne la morte. Infine, gli anticorpi possono attivare il sistema del complemento, una famiglia di proteine ed enzimi proteolitici, sia di membrana che solubili: esse in ultimo formano un complesso finale che le opsonizza oppure s’inserisce nella membrana delle cellule bersaglio, causando lisi osmotica.
Fonti:
Abbas, Lichtman, “Fondamenti di immunologia”, Piccin