I cromosomi sono costituiti da cromatina superavvolta su un probabile scaffold proteico (cfr risposta). Durante la mitosi, cioè la generazione di due cellule figlie distinte a partire da una cellula madre, i cromosomi hanno il fondamentale compito di permettere la suddivisione esatta del materiale genetico presente nella cellula originaria tra le due cellule figlie.
I meccanismi che regolano l’inizio della mitosi e quindi la formazione dei cromosomi sono ancora ben lontani dall’essere completamente compresi, tuttavia numerosi passi avanti sono stati effettuati in questo campo grazie alla biologia molecolare, allo studio delle cellule in vitro e alle nuove tecniche di microscopia ultrastrutturale.
Una cellula classica segue un ciclo vitale costituito da due grandi fasi: la mitosi e l’interfase. L’interfase è a sua volta costituita da tre momenti, la fase G1, la fase S e la fase G2. La lunghezza delle singole fasi non è sempre costante e dipende dal tipo di cellula preso in considerazione e il passaggio da una fase alla successiva è regolato da complessi meccanismi che tengono conto sia di un orologio interno che dei segnali provenienti dall’ambiente extracellulare (ormoni, matrice, tessuto).
Uno dei fattori studiati coinvolti nella temporizzazione del ciclo cellulare è il sistema delle cicline-cdk chinasi. Queste proteine presentano un’ampia variazione di concentrazione nelle diverse fasi del ciclo cellulare. Ad esempio, il complesso ciclina cdk1-B1, che fosforila numerosi substrati, presenta un’onda di concentrazione proprio in concomitanza con la mitosi e sembra determinare il passaggio dalla fase G2 alla mitosi.
Tra i substrati fosforilati da questo complesso ci sono:
– proteine associate all’assemblamento del fuso mitotico quali laminine nucleari, componenti della matrice del Golgi, proteine motrici e MAP (proteine associate ai microtubuli) e il fattore che promuove l’anafase
– molecole coinvolte nella condensazione dei cromosomi quali la proteina passeggera Aurora B chinasi, i complessi multiproteici delle condensine e quelli delle coesine
– altre chinasi mitotiche come polo-like chinasi che si ritrova a livello dei centrosomi.
Un fattore importante per la formazione dei cromosomi mitotici è la fosforilazione delle code N terminali degli istoni che formano il nucleosoma, soprattutto dell’istone H3, che viene fosforilato dalla proteina passeggera Arora B chinasi.
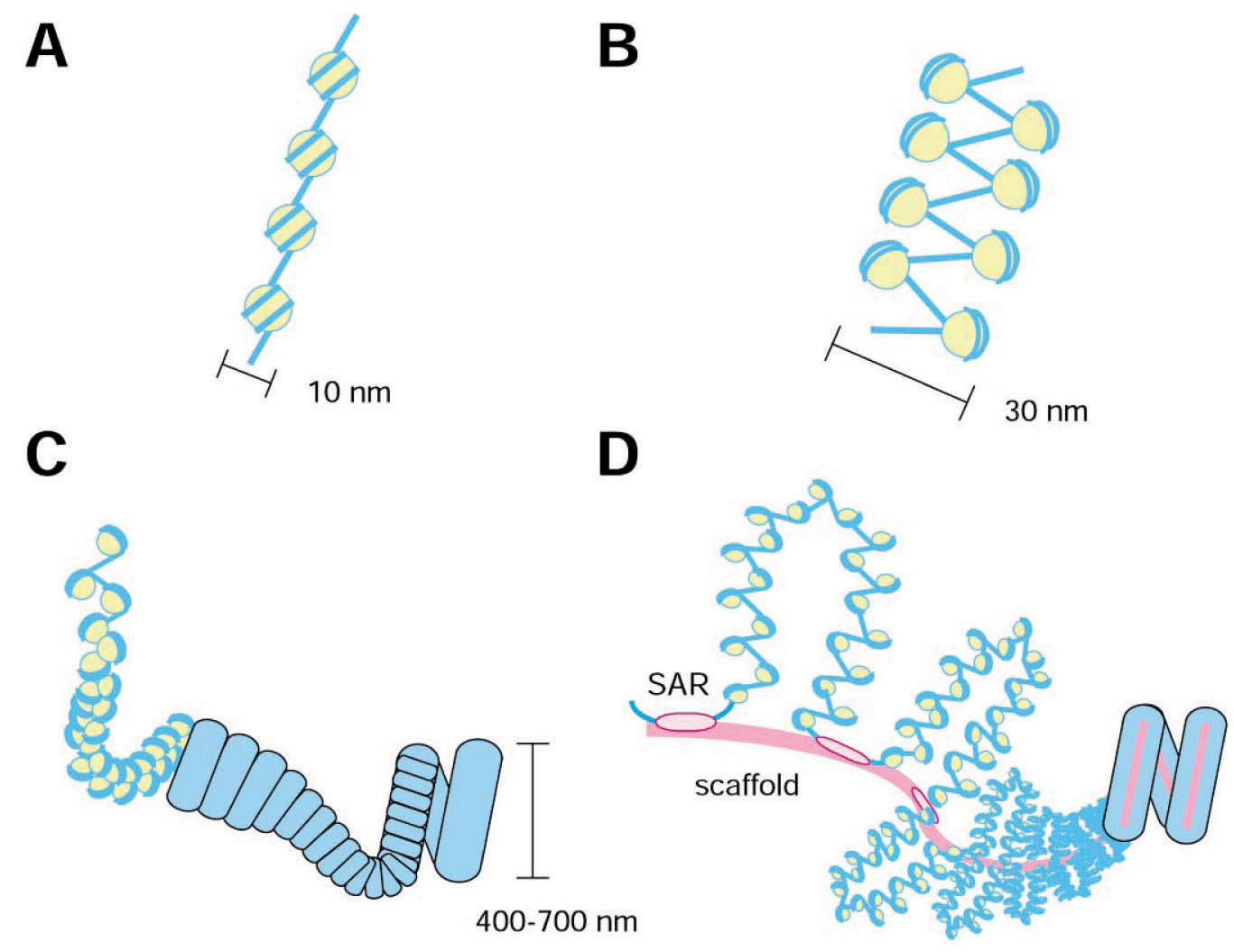
Modello gerarchico per l’organizzazione cromosomica: A) fibra nucleosomica. B) fibra cromatinica a 30 nm. C) Fibre di ordine superiore. D) organizzazione delle fibre di ordine superiore attorno allo scaffold proteico. (da Swedlow and Hirano, 2003)
L’unità fondamentale dell’organizzazione cromatinica è la fibra nucleosomica, costituita da DNA a doppio filamento arrotolato attorno a un ottametro di istoni, che permette una compattazione di 6-7 volte la lunghezza del DNA. La fibra cromatinica, di ordine superiore, è formata da una fibra incrociata di circa 30 nm, tuttavia ci sono state numerose controversie al riguardo della descrizione di questa struttura, la fibra a 30 nm è solitamente vista come un ripiegamento o un avvolgimento ad elica delle fibre nucleosomiche e forma un’ulteriore compattazione di 6-7 volte. Allo stadio di mitosi, le fibre si possono compattare ulteriormente di 200-500 volte, raggiungendo una compattazione lineare finale di 10.000-20.000 volte nel cromosoma mitotico. Questi livelli di assemblamento cromatinico, cosiddetti superiori, sono delicati quanto controversi. Una visione classica suggerisce il ripiegamento o l’avvolgimento ad elica delle fibre a 30 nm in strutture sempre più ampie. E’ stato proposto che la cromatina in metafase sia organizzara in anse attaccate a una struttura sottostante ricca di cromatina chiamata scaffold metafasico. Questo scaffold è stato visualizzato tramite microscopia elettronica in cromosomi privati degli istoni; il risultato mostra anse di dna di circa 40 kb che si dipartono da un aggregato di proteine che ha la forma del cromosoma originale. Gli scaffold isolati contengono proteine non istoniche, inclusa la topoisomerasi II. Lo scaffold potrebbe essere sia un asse, presumibilmente costituito da DNA e/o proteine che servono come scheletro del cromosoma, o una rete lassa di complessi DNA/proteine che regolano l’assemblamento della struttura a livello superiore dei cromosomi.
Generalmente i modelli dello scaffold prevedono la presenza di specifici fattori proteici e di sequenze cis di DNA che localizzano i limiti di queste unità e definiscono il loro attacco come principali organizzatori della cromatina. Nella maggior parte dei modelli,due elementi vicini, sebbene separati fino a 100kb di DNA, si legano allo scaffold con l’intervento del DNA che forma un’ansa. Una sequenza di DNA che è stata proposta funzionare come sequenza cis nell’assemblamento del cromosoma ciene chiamata la regione di attacco allo scaffold (SAR). Questa sequenza moltoricca di AT potrebbe agire come sito di legame per la DNA topoisomerasi II, una componente dello scaffold cromosomico. In alcune condizioni sperimentali è stato visualizzato nei cromosomi mitotici un assemblamento elicoidale, suggerendo che l’organizzazione di ordine superiore del cromosoma sia un arrangiamento elicoidale delle braccia cromosomiche.
In questi link si trovano alcune indicazioni sull’organizzazione spaziale dei cromosomi e sulla loro formazione in mitosi.