Con il termine sauropsidi si usa indicare rettili e uccelli, entrambi possiedono uova ricche di tuorlo. Il più conosciuto e studiato tra i Sauropsidi è sicuramente il pollo, quindi utilizzeremo questo modello per spiegare gli eventi che portano alla formazione dei tre foglietti embrionali. I Sauropsidi presentano una segmentazione meroblastica discoidale che da origine a un blastodisco posizionato sulla massa del tuorlo. Le cellule del blastodisco ingredono o delaminano dall’epiblasto esterno nella cavità sottogerminale a formare lo strato dell’ipoblasto primario. In un secondo momento, le cellule che formano il margine posteriore si spostano anteriormente fino a raggiungere le cellule dell’ipoblasto primario e prendono il nome di ipoblasto secondario. L’epiblasto e l’ipoblasto sono uniti insieme lungo il margine e lo spazio creatosi tra di loro si chiama blastocele. L’ipoblasto non contribuisce alla formazione dell’embrione ma formerà le membrane che circondano il sacco del tuorlo.
Attraverso il solco primitivo, formatosi lungo l’asse antero-posteriore dell’embrione, ingredono per prime le cellule che formeranno l’endoderma dell’intestino, anche se non avviene la formazione di un archenteron primitivo. Una volta entrate nel blastocele, queste cellule migrano anteriormente e successivamente sostituiscono le cellule dell’ipoblasto nella porzione anteriore dell’embrione. Mentre la stria primitiva si sposta in avanti, anche le cellule dell’ipoblasto secondario migrano anteriormente dal margine posteriore del blastodisco. Il secondo gruppo di cellule che entra nel blastocele attraverso il nodo di Hensen, si spostano anteriormente ma restano tra l’endoderma e l’epiblasto a formare il mesoderma della testa e il cordomesoderma. Le altre porzioni di endoderma e mesoderma vengono invece formate dalle cellule che ingredono attraverso le porzioni laterali della stria primitiva. Queste cellule si dividono in due gruppi, uno dei quali si muove più in profondità e spinge lateralmente le cellule dell’ipoblasto. Questo gruppo da origine all’endoderma e a molti annessi extraembrionali. Il secondo gruppo migra nel blastocele tra l’epiblasto e l’ipoblasto e forma un foglietto lasso di mesenchima che originerà le strutture mesodermiche.

Ricordiamo in particolare che il mesoderma laterale si divide in una somatopleura addossata all’ectoderma e una splancnopleura addossata all’endoderma.
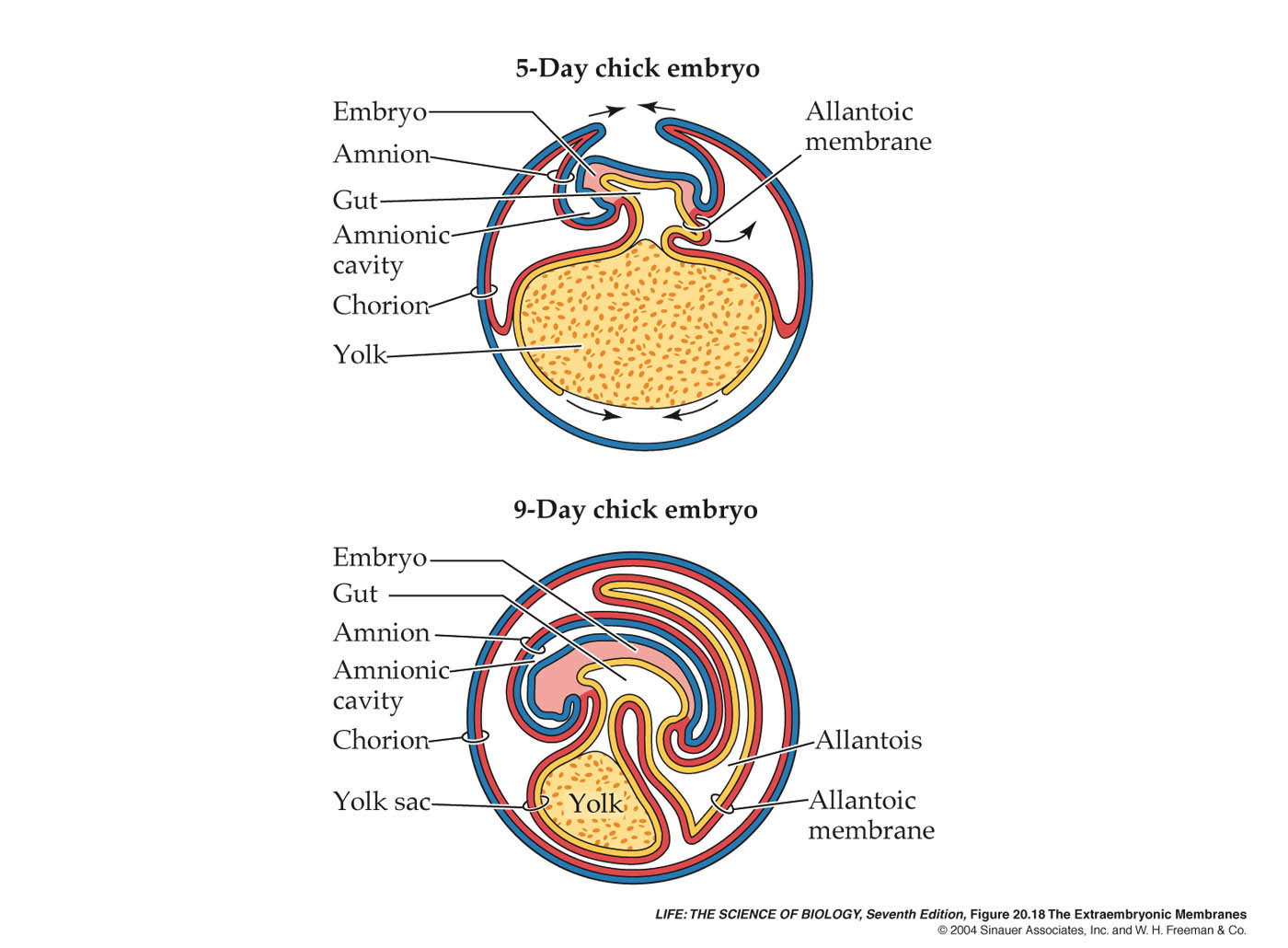
Per quanto riguarda gli annessi embrionali, ricordiamo che nei sauropsidi ritroviamo: il sacco vitellino, il corion, l’allantoide e l’amnios. Ognuna di queste strutture ha una precisa funzione, indispensabile per lo sviluppo dell’embrione. Il sacco del tuorlo ha il compito di demolire e metabolizzare il tuorlo, permettendo il trasferimento dei suoi costituenti all’embrione. L’amnios secerne e mantiene il liquido amniotico, una soluzione acquosa che protegge l’embrione dalla disidratazione e che ha permesso ai sauropsidi di conquistare la terra ferma svincolandosi dall’ambiente acquatico per la riproduzione. L’allantoide è connesso con l’intestino il formazione e costituisce un organo sacciforme atto al deposito dei materiali di rifiuto, quali i composti azotati, inoltre l’allantoide svolge un importantissima funzione nel permettere gli scambi gassosi attraverso il guscio poroso e si accresce con il procedere dello sviluppo, così come diminuisce il sacco del tuorlo. il corion è l’annesso più esterno, riveste tutti gli altri e media gli scambi metabolici tra i territori embrionali e il mondo esterno.
Andando nello specifico della loro formazione, il sacco vitellino si origina da epibolia dei tre foglietti embrionali, che si spostano verso il polo vegetativo ricoprendo quasi completamente il tuorlo. L’ectoderma extraembrionale che è derivato dall’ipoblasto, si accresce e trasporta con se la porzione somatopleurica del mesoderma. Parimenti l’endoderma trascina con se quella splancnoplerurica. La cavità così formatasi viene chiamata celoma extraembrionale. Quasi alla fine dell’epibolia, la parete esterna (ectoderma e somatopleura) si solleva a livello dell’estremità cefalica dell’embrione, formando una plica che viene chiamata proamnios. Le pliche proamniotiche sono due, una per lato, e si incontrano, saldandosi tra di loro a formare l’amnios, o meglio il pleuramnios, e richiudendo l’embrione in una cavità detta amniotica. Questa cavità ben presto si riempie di un liquido secreto dalle cellule della parete. Il sollevamento e la fusione delle pliche dilata il celoma extraembrionale, in tal modo la parete interna (endoderma e splancnopleura) vanno a costituire il sacco vitellino rivestendo completamente il tuorlo. La parete esterna invece va a circondare l’intero embrione e gli annessi precedentemente citati, e prende il nome di corion. Il corion esternamente si trova in contatto con l’albume che circonda l’uovo. Infine, posteriormente al sacco vitellino, compare dall’intestino una piega. Questa piega si espande nel celoma extraembrionale fino a toccare la faccia interna del corion. Quest’ultimo annesso è l’allantoide e la sua unione con il corion dà origine alla membrana corioallantoidea. Nell’allantoide si depositano cristalli di acido urico, che grazie alla loro igroscopicità assorbono una gran quantità di acqua. L’allantoide negli uccelli si espande sotto il sacco del tuorlo (rimasto aperto a livello dell’ombelico vitellino, e unito al corion forma delle espansioni villose che hanno la funzione di assorbire acqua e albumine dall’albume. Questa nuova struttura si chiama appunto sacco dell’albume.
Per una visione generale dei processi di gastrulazione: http://www.vialattea.net/esperti/php/risposta.php?num=4083
Per approfondire i processi di formazione della stria primitiva e della notocorda: http://www.vialattea.net/esperti/php/risposta.php?num=8303