Molti uccelli hanno in comune gli stessi meccanismi di orientamento spaziale, indipendentemente dalle loro prestazioni migratorie più o meno spiccate e dai gruppi sistematici di appartenenza. Le capacità di orientamento nello spazio comprendono sia meccanismi di vera navigazione, che meccanismi di mantenimento e scelta di direzioni mediate dalla lettura di "bussole" basate sulla percezione di svariati input sensoriali.
Queste bussole, basate sia su componenti innate che sull’apprendimento programmato, sono meccanismi che consentono agli esseri viventi di mantenere direzioni costanti sfruttando sistemi di riferimento esterni, quali gli astri, il sole, il sole come fonte di luce polarizzata ed il campo geomagnetico.
In particolare la bussola magnetica è un meccanismo di orientamento molto diffuso nel regno animale, il suo utilizzo è stato dimostrato nei Passeriformi, nelle api, nel colombo, nei roditori, nei talitri etc, svolgendo un ruolo molto importante in tutte quelle condizioni che impediscono l’utilizzo del Sole (ad esempio di notte, a cielo coperto, sotto terra).
Gli esperimenti più significativi sull’orientamento magnetico sono stati condotti sui Passeriformi migratori notturni. L’utilizzo del campo geomagnetico da parte degli uccelli migratori cominciò ad essere ipotizzato partendo dall’osservazione che essi erano orientati anche quando, per la copertura del cielo, nè il Sole e nè le stelle erano utilizzabili. Seguendo questa traccia W. e R. Wiltschko dimostrarono nel 1968 come l’orientamento di pettirossi ingabbiati e saggiati nel periodo migratorio poteva essere influenzato dalla presenza di campi magnetici artificiali: gli uccelli rispondevano ad una rotazione del nord magnetico ottenuta tramite spirali di Helmholtz, con una corrispondente rotazione nella preferenza direzionale, che corrispondeva sia qualitativamente che quantitativamente al trattamento effettuato, dimostrando così che essi utilizzano il campo magnetico come riferimento direzionale.
Fondamentalmente per rilevare la capacità di percepire il campo magnetico, vengono effettuati due tipi di esperimenti: gli "esperimenti di condizionamento a stimoli magnetici" e gli "esperimenti di orientamento magnetico". Nel primo tipo di esperimenti si cerca di indurre una risposta differenziale (in genere una scelta legata all’ottenimento di cibo) a seconda della presenza o meno di un campo magnetico artificiale. Questo approccio è stato tentato più volte, ma con buoni risultati solo nelle api. Nel secondo tipo di esperimenti come procedura generale si va invece a indurre una variazione nel campo magnetico terrestre attorno alla zona in cui si ha orientamento e si tenta di rilevare le conseguenti risposte nell’animale. Il metodo più semplice, ma più generico, consiste nell’applicazione di magneti, in genere mobili, con l’attesa di indurre un disorientamento se gli animali si basano su stimoli magnetici. Un sistema più preciso, ma usabile solo in condizioni controllate, consiste nell’uso di bobine magnetiche (spirali di Helmholtz o cubo di Rubens), con cui produrre specifici campi magnetici alterati, determinabili con precisione nel’area di studio.
Alcuni animali come ad esempio i salmoni, o i ratti-talpa sembrano in grado di riconoscere la polarità del campo magnetico, mentre nella maggior parte dei casi la bussola funziona come una bussola ad inclinazione: l’animale non appare capace di distinguere il senso delle linee di forza del campo magnetico, ma solo la loro inclinazione rispetto alla superficie terrestre e ciò fornirebbe loro delle informazioni riguardo la propria posizione rispetto alla direzione "verso il polo" e "verso l’equatore", senza dare informazioni sulla polarità. Ciò è stato dimostrato sempre nei pettirossi andandoli a saggiare in un campo magnetico con la componente verticale invertita.
Per quanto riguarda l’organo di senso implicato nella percezione dello stimolo magnetico negli uccelli, sappiamo oramai da tempo che anche stimoli e recettori lontani dal nostro bagaglio sensoriale possono giocare un ruolo cruciale nel guidare gli animali, ma le nostre conoscenze sui meccanismi fisiologici e neurobiologici che stanno alla base di questa magnetorecezione sono ancora piuttosto limitate. Per una completa comprensione del cosiddetto "senso magnetico" è necessario conoscere la natura e la localizzazione dei recettori, i meccanismi di trasduzione coinvolti, le connessioni col SNC e la parte del cervello implicata nell’elaborazione dell’informazione magnetica.
Solo di recente sono stati proposti due possibili meccanismi di trasduzione per spiegare la magnetorecezione:
– PERCEZIONE TRAMITE PARTICELLE FERROMAGNETICHE nel colombo sono state rinvenute all’interno delle terminazioni nervose del ramo oftalmico mediale, che è una branca del nervo trigemino,
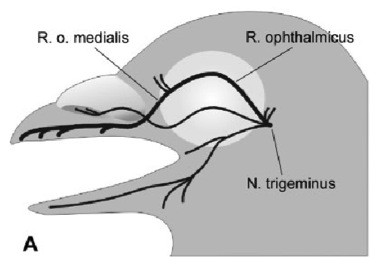
delle formazioni, che si è ipotizzato siano implicate nella rivelazione di stimoli geomagnetici, utili per il meccanismo di mappa.

Tali formazioni sono costituite da un particolare tipo di magnetite biogenica, detta "superparamagnetica", che in presenza di un campo magnetico esterno si polarizza, con conseguenti interazioni dipolo che portano a seconda della direzione del campo ad una deformazione della membrana a cui sono legate ed ad una apertura di canali ionici meccano-sensibili, che vanno ad influire sul potenziale di membrana.
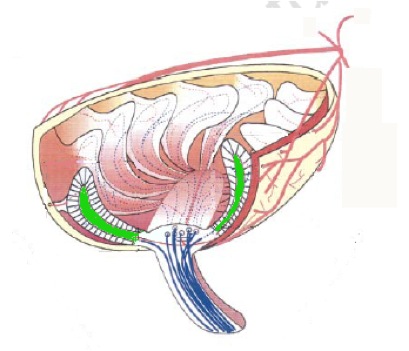
Nella trota iridea invece sono state rinvenute particelle di magnetite "a dominio singolo" (indicate in verde nella figura sottostante) all’interno della lamina basale dell’epitelio olfattivo, il quale è contenuto nella capsula nasale, che è innervata da una branca del nervo trigemino (ros V).
– PERCEZIONE CHIMICA: IL "RADICAL PAIR MODEL" questo modello prevede che il decorso ed i prodotti di reazioni chimiche che comportano la formazione di radicali liberi possano dipendere da campi deboli come il campo geomagnetico e che questo determinerebbe una prima trasduzione dello stimolo magnetico. In particolare si ritiene che il campo magnetico influenzi la dinamica di interconversione tra i due stati in cui si può venire a trovare la coppia radicalica e quindi il tutto dipenderebbe dall’allineamento che la coppia assume rispetto al campo magnetico locale. Attualmente l’ipotesi più accreditata è quella basata su un radical pair model che avviene nelle cellule fotorecettoriali retiniche, a carico di fotopigmenti retinici chiamati "criptocromi", che sono le uniche molecole fotoeccitabili presenti nella retina dei Vertebrati, che possono soddisfare i requisiti previsti dal modello. I prodotti di queste reazioni andrebbero poi ad influenzare le vie di trasduzione visive, portando così ad una percezione del campo magnetico sottoforma di diversi patterns visivi, in base ai diversi orientamenti della testa, utile solo per il meccanismo di bussola ad inclinazione.
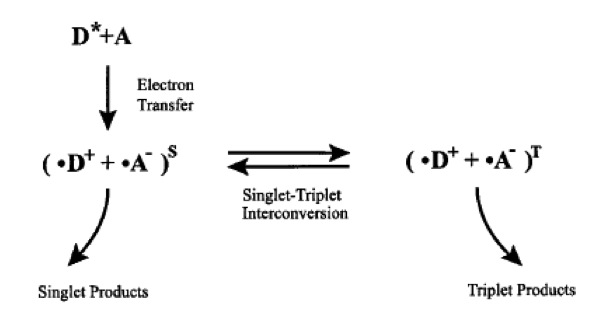
Indicazioni a favore di questo modello derivano da diversi tipi di esperimenti comportamentali, i quali hanno indicato un possibile link tra magnetorecezione e sistema visivo. La maggior parte delle evidenze derivano da esperimenti di orientamento con luci a diversa lunghezza d’onda, effettuati inzialmente nel tritone Notophtalmus e poi replicati ampiamente in Drosophila, in Tenebrio, nei topi e soprattutto in vari Passeriformi migratori. Ulteriori prove derivano poi da esperimenti con campi oscillanti a radio frequenza.
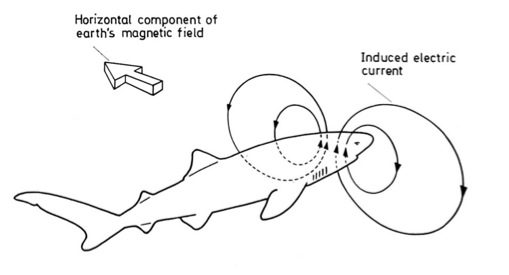
– PERCEZIONE PER INDUZIONE in questo le informazioni magnetiche sono percepite da elettrocettori, che rilevano il campo elettrico indotto dal movimento dell’animale in un campo magnetico. Il campo elettrico indotto dipenderebbe dalla direzione e dall’intensità del campo magnetico attraversato e dal rapporto spaziale tra la direzione del movimento e quella del campo magnetico terrestre. Tutto ciò è in effetti ciò che si ritiene avvenga nelle ampolle del Lorenzini degli Elasmobranchi, ma ci sono scarse prove sperimentali che effettivamente utilizzino gli elettrocettori per derivare informazioni direzionali.
Per ulteriori approfondimenti:
Wiltschko R, Wiltschko W (1995). "Magnetic orientation in animals". Springer, Berlin, Heildeberg, New York.
Liedvogel M, Mouritsen H (2009). "Cryptochromes-a potential magnetoreceptor: what do we know and what do we want to know?". J. R. Soc. Interface doi:10.1098/rsif.2009.0411.focus (interessante e aggiornato review sui criptocromi coinvolti nella percezione chimica).
Cadiou H, McNaughton P. A (2009). "Avian magnetite-based magnetoreception: a physiologist’s perspective". J. R. Soc. Interface doi:10.1098/rsif.2009.0423.focus (interessante e aggiornato review sul modello basato sulle particelle di magnetite).

Balia nera (Ficedula hypoleuca)