La parete dei batteri Gram negativi è caratterizzata dalla presenza di un sottile strato di peptidoglicano con pochi legami crociati.
Sono presenti anche altri componenti oltre al pepidoglicano che si associano a formare una struttura esterna allo strato mureinico detto membrana esterna. 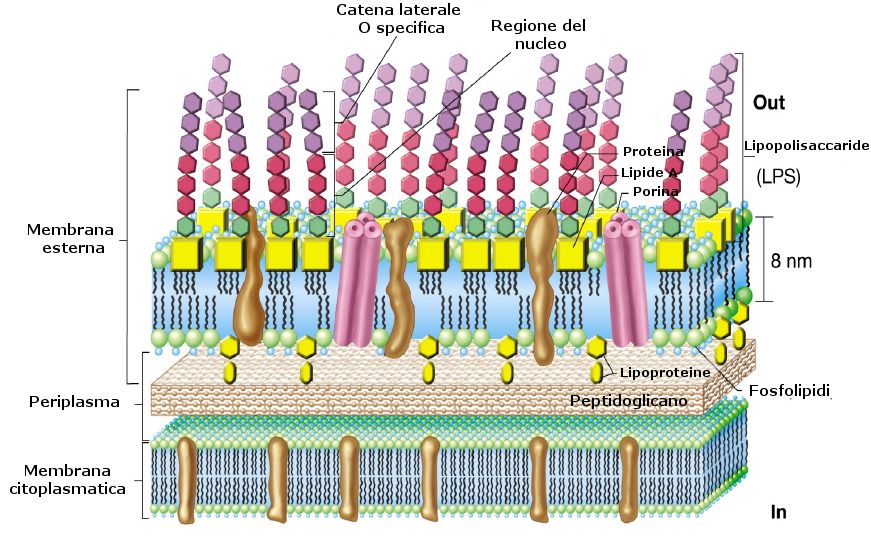
La membrana esterna ha la struttura a due foglietti tipica di tutte le membrane biologiche.
Immersi in tale struttura vi sono diversi componenti:
Fosfolipidi: simili a quelli della membrana citoplasmatica e distribuiti sul foglietto interno
Proteine: inserite a varie altezze
Lipoproteine: hanno funzione di ancoraggio della membrana esterna allo strato sottostante di peptidoglicano
Lipolisaccaride: costituisce con la sua componente lipidica la maggior parte del foglietto esterno ed è il principale responsabile della tossicità batterica.
Struttura del lipolisaccaride
Ha una porzione polisaccaridica rivolta verso l’esterno ed una porzione lipidica immersa nel foglietto esterno della membrana esterna.
Lo si può suddividere in tre parti, dall’esterno verso l’interno:
Catena laterale O-specifica (in viola nella figura seguente): è un polimero costituito da unità oligosaccaridiche di ripetizione (in media 40 unità) e vi si possono ritrovare almeno 20 zuccheri differenti. Rappresenta la parte più variabile della classificazione sierologia dei batteri Gram- .
Regione del nucleo (core):
è un oligodisaccaride costante nell’ambito di un genere. Lo si può suddividere in Outer core (in fucsia) costituito da unità variabili di zuccheri a 6 atomi di carbonio ed inner core (in verde) costituito in tutti i batteri gram –da unità di uno zucchero ad 8 atomi di carbonio, il 2-keto-3-deoxioctulosio.
Lipide A (in giallo):
è un glicofosfolipide costituito da un dimero di N-acetilglucosammina fosforilata avente legati degli acidi grassi a catena lunga. Rappresenta la porzione idrofoba che si ancora alla membrana esterna. Rappresenta il principio tossico dell’intera molecola ed è altamente conservato in tutti i batteri Gram -.
Struttura del lipide A
Come si può notare dalla figura i due monomeri di N-acetilglucosammina hanno tutte le funzioni alcoliche sostituite (fosforilate o esterificate con acidi grassi a lunga catena). Si prenda in considerazione il carbonio in posizione 1 in entrambi gli zuccheri (detto anche carbonio anomerico):
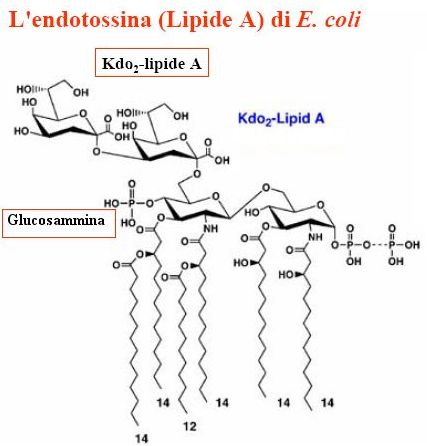
- nel caso dello zucchero di sinistra questo è “bloccato” nella formazione del legame β-glicosidico con il secondo monomero di glucosammina;
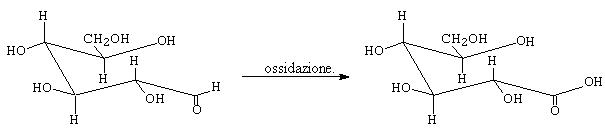
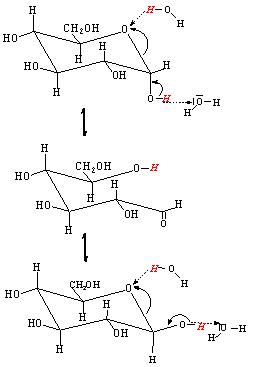
- lo zucchero di destra ha il carbonio anomerico fosforilato. Tale fosfato può però staccarsi e ridare un gruppo OH libero e reattivo. Tale gruppo può andare incontro a reazione di mutarotazione (figura seguente) dando la forma aperta con gruppo funzionale aldeidico libero. L’aldeide può a sua volta ossidarsi (ultima figura) ad acido carbossilico in presenza di particolari reagenti che vengono ridotti e pertanto gli zuccheri che danno tali reazioni sono detti “riducenti”. Alcuni saggi qualitativi (Tollens, Feheling) , specifici per le aldeidi, sono utilizzati per evidenziare gli zuccheri riducenti sfruttando proprio questa specifica reattività.
Esempio di mutarotazione con apertura del ciclo nel caso dell’α D-glucopiranosio
Esempio di blanda ossidazione che agisce solo sulle funzioni aldeidiche e non su quelle alcoliche (si passa da aldeide ad acido carbossilico).